Часы с Wi-Fi на газоразрядных индикаторах ИН-12 или ещё один тёплый ламповый свет
Здравствуйте, уважаемые читатели. Тема электронных часов меня никак не отпускает, и сегодня будет рассказ об очередном проекте. Я уже делал часы на газоразрядных индикаторах, ознакомиться можно здесь. Новые часы также будут на советских лампах ИН-12, но немного сложнее — с Wi-Fi подключением к сети, синхронизацией времени по NTP, со схемой часов реального времени и управлением через веб-интерфейс.
Под катом — схема, печатная плата, прошивка, процесс сборки корпуса, описание функционала часов.
Попалось мне как-то на YouTube видео с этими часами, автором данных часов является Константин Репников. Оказалось что печатные платы для данного проекта автор выложил на PCBWay, это было решающим фактором, решил заказать платы и собрать себе такие часы.
Схема
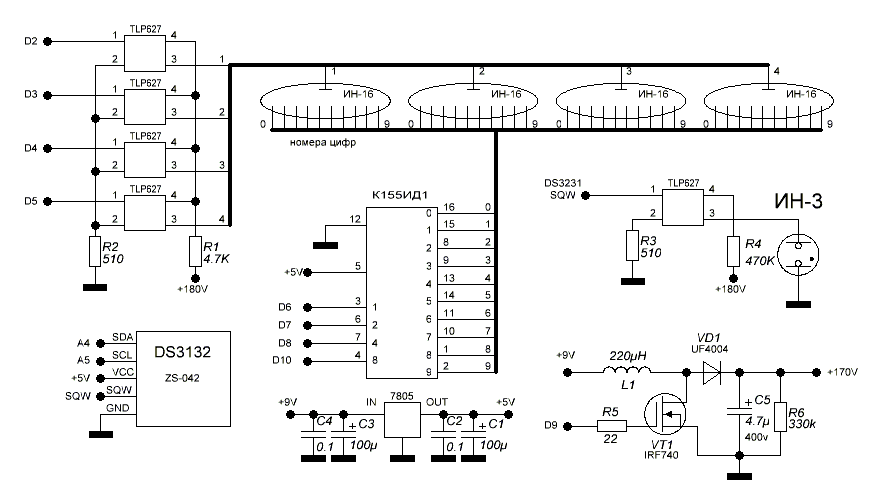
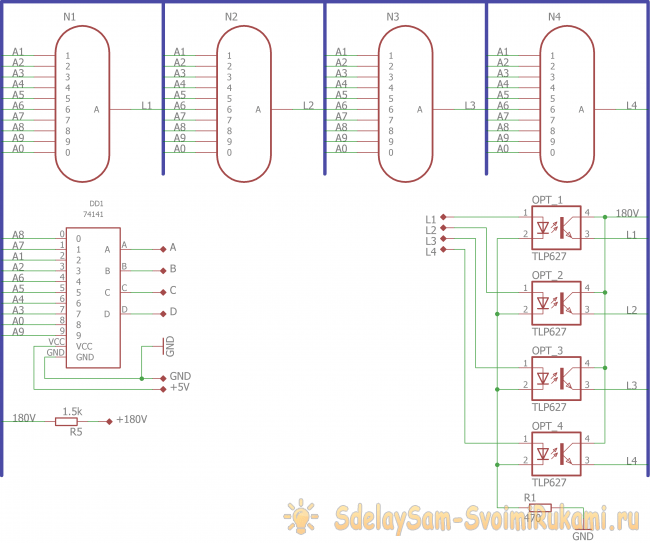
Устройство собрано на основе микроконтроллера ESP8266, это позволяет реализовать управление настройками часов через браузер, и синхронизацию времени с помощью NTP протокола, схема устройства показана на рисунке ниже:
 Напряжение 3,3 В для питания микроконтроллера обеспечивает DC-DC преобразователь на микросхеме MC34063AD. Высокое напряжение 170 В для питания ламп реализовано с помощью на повышающего преобразователя на микросхеме MAX1771. Также в схеме имеются часы реального времени на микросхеме DS3231. Питается схема от внешнего источника напряжения 9 — 12 В.
Напряжение 3,3 В для питания микроконтроллера обеспечивает DC-DC преобразователь на микросхеме MC34063AD. Высокое напряжение 170 В для питания ламп реализовано с помощью на повышающего преобразователя на микросхеме MAX1771. Также в схеме имеются часы реального времени на микросхеме DS3231. Питается схема от внешнего источника напряжения 9 — 12 В.Печатные платы это набор из трех плат с двухсторонним монтажом, вид с обеих сторон:
Платы выполнены довольно качественно — паяльная маска (я выбрал чёрного цвета), металлизация отверстий, шелкография все дела.
Сборка
Пока ждал платы, заказал также у китайцев детали для сборки, список всех необходимых компонентов со ссылками под спойлером.
Список деталей для сборки
• DS3231N SOIC-16W — 1 шт.
• ESP-12F — 1 шт.
• 74HC595, 74HCT595 SOIC-16 — 2 шт.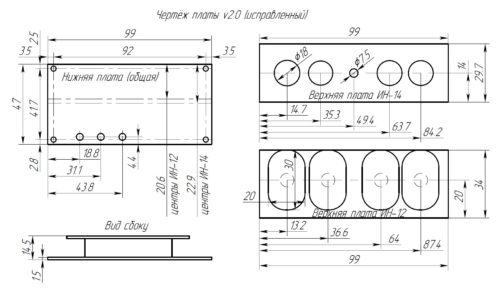
• MAX1771ESA SOIC-8 — 1 шт.
• MC34063AD SOIC-8 — 1 шт.
• IRF740, IRF840, TO-220-3 — 1 шт.
• 220uH L_12x12mm_H6mm — 2 шт.
• US1M (BA159) 300V 1A SMA (DO-214AC) — 1 шт.
• SS24, SMA (DO-214AC) — 1 шт.
• 0.1uF 0805 — 9 шт.
• 470pF 0805 — 1 шт.
• 100uF 6.3V D6.3mm_P2.50mm — 4 шт.
• 3.3uF — 4.7uF 200V D10.0mm_P5.00mm — 1 шт.
• 220uF 16V D6.3mm_P2.50mm — 4 шт.
• 1ом 0805 1206 — 8 шт.
• 1k 0805 — 1 шт.
• 2k 0805 — 1 шт.
• 3.3k 0805 — 18 шт.
• 10k 0805 — 14 шт.
• 1m 0805 — 6 шт.
• 18k 0805 — 1 шт.
• R PHOTO 5537 — 1 шт.
• MMBTA42 SOT-23 — 15 шт.
• MMBTA92 SOT-23 — 4 шт.
• 5,5 x 2,1 connector — 1 шт.
• Neon light bulb lamp — 2 шт.
• ИН-12 — 4 шт. (ищите по месту, у китайцев дорого)
Если кто также захочет собрать эти часы, стоит сказать, что некоторые детали достаточно мелкие, для многих это может быть проблема. Для понимания фото транзистора на пальце:
Для понимания фото транзистора на пальце:
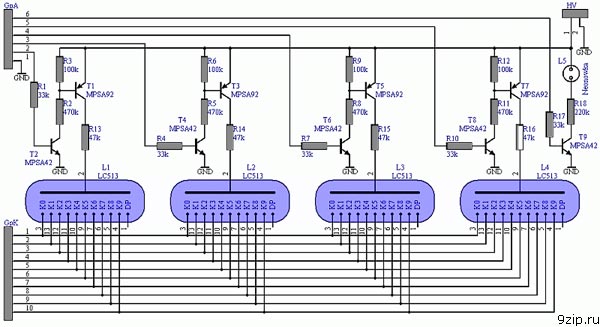
Если не брать во внимание что детали маленькие, собирать часы довольно просто, где что должно быть всё подписано — попутать тяжело. После нескольких дней с паяльником и лупой, разрезания дремелем и отмывания спиртом, имеем три платы. Первую я условно назвал «блок питания», вид с обеих сторон:
Вторая плата — плата часов:
Третья плата — плата индикации:
У неонок, которые служат двоеточием между цифрами, на каждый вывод одел тонкую термоусадку, после вставил в отрезок черной оболочки от кабеля. Все три платы соединяются вместе в «сэндвич» с помощью PLS-PLD разъёмов, получается такая конструкция:
Прошивка
Чтобы часы заработали нужно прошить микроконтроллер, я использовал популярный программатор Ch441A. Соединяем плату часов с программатором по схеме на фото:
Перемычку на программаторе переставляем из положения 1-2 в положение 2-3, тем самым переключаем его в режим USB — UART конвертера.
 Подключаем всё это дело к компьютеру, в диспетчере устройств должен появиться новый виртуальный COM порт:
Подключаем всё это дело к компьютеру, в диспетчере устройств должен появиться новый виртуальный COM порт:Для прошивки ESP8266 нам понадобится программа NodeMCU Flasher, запускаем файл ESP8266Flasher.exe. В первой вкладке «Operation» выбираем нужный COM порт (у меня выбрался автоматически):
Идём во вкладку «Config», рядом с зеленой строкой с надписью «INTERNAL://NODEMCU» жмём на шестеренку и выбираем прошивку в формате *.bin, так же выбираем «0x00000» как показано на рисунке ниже:
Далее переходим во вкладку «Advanced» и меняем там скорость на 115200:
Возвращаемся на вкладку «Operation» и жмём «Flash»:
Начнется прошивка контроллера, по окончании в нижнем левом углу должна появиться галочка в зеленом кружочке:
Ссылка на файл прошивки на Google Диске — тут
Внимание! В схеме присутствует опасное для жизни высокое напряжение, настоятельно рекомендуется быть предельно осторожным!
Собираем часы и подключаем питание, часы должны показать полночь (засветятся нули):
Открываем на смартфоне настройки Wi-Fi, в списке должна появиться новая точка доступа «NixieIN12», подключаемся к ней:
После подключения откроется веб-интерфейс часов, по умолчанию откроется раздел «WiFi settings» пока нам это не нужно, выбираем «Time settings» и жмем кнопку «Set time»:
Часы синхронизируются и покажут текущее время:
Электронная часть работает, надо делать корпус.

Корпус
Корпус как у автора мне не понравился, какой-то он «чрезмерно круглый», решил сделать свой. Корпус будет из фанеры толщиной 4 мм, вырезаться лазером на ЧПУ станке. Нарисовал чертеж с деталями корпуса в CorelDRAW:
После лазерной резки, получил такой комплект фанерок:
Намазываем детали клеем, аккуратно собираем и зажимаем в тисках на сутки сохнуть:
После склейки получилось следующее:
Зачищаем снаружи корпус наждачной бумагой, чтобы сгладить все неровности и удалить нагар после лазерной резки, внутренности я решил оставить обожженными, после обрабатываем льняным маслом:
С тыльной стороны:
К днищу прикрепил саморезами две резиновые ножки:
Вставляем батарейку 2032, пластиковыми стойками прикрепляем заднюю стенку корпуса:
Собираем всё вместе:
Функционал
Как упоминалось выше, при первом включении часы создают точку доступа «NixieIN12», после подключения к ней откроется веб-интерфейс часов с разделом «WiFi settings».
http://nixiein12
Либо ввести ip-адрес который выдал нашим часам роутер (можно посмотреть в веб-интерфейсе роутера). Должно открыться такое меню веб-интерфейса:
Все разделы веб-интерфейса в развёрнутом виде:
Здесь у нас и установка времени, настройка будильников, настройка оповещения по времени, настройки Wi-Fi, настройки часов, сервисное меню для настроек антиотравления катодов, настройки светодиодной подсветки, и наконец предустановки яркости свечения ламп.
В часах имеется три будильника и оповещения (например каждый час) при условии подключения модуля DFPlayer mini:
Есть также возможность подсвечивать лампы светодиодами WS2812B, на плате часов есть разъём для подключения — в данном экземпляре часов это не реализовано, мне такое не нравится, теряется вся «ламповость».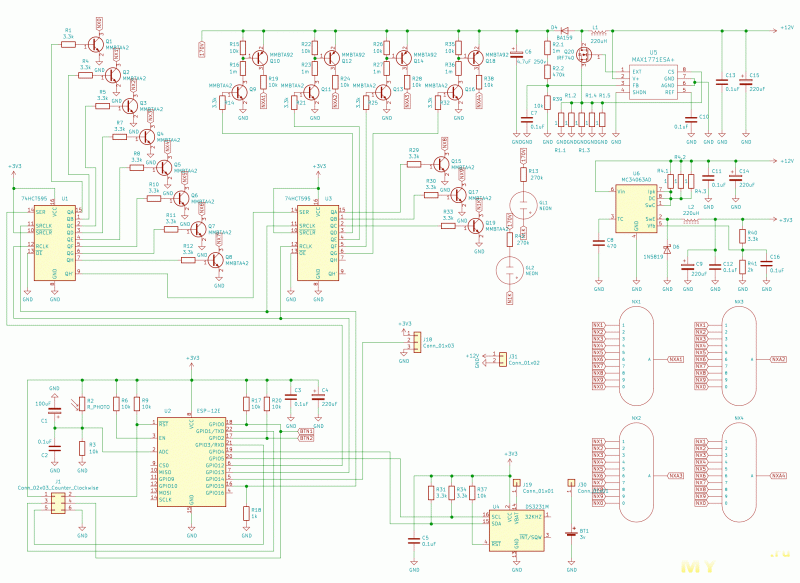
Стоит также показать как меняются цифры на «дисплее», они плавно затухают и плавно зажигаются:
Под конец ещё пару фото как выглядит эта красота в темноте:
Часы работают уже около месяца, при обесточивании и последующем включении время не сбрасывается, схема часов реального времени работает. Обратил внимание что всегда показывают время одинаковое с моим смартфоном, т. е. синхронизация также работает исправно.
В итоге имеем ещё одну красивую вещь, сделанную своими руками.
Всем удачных поделок, спасибо за внимание, и будьте здоровы.
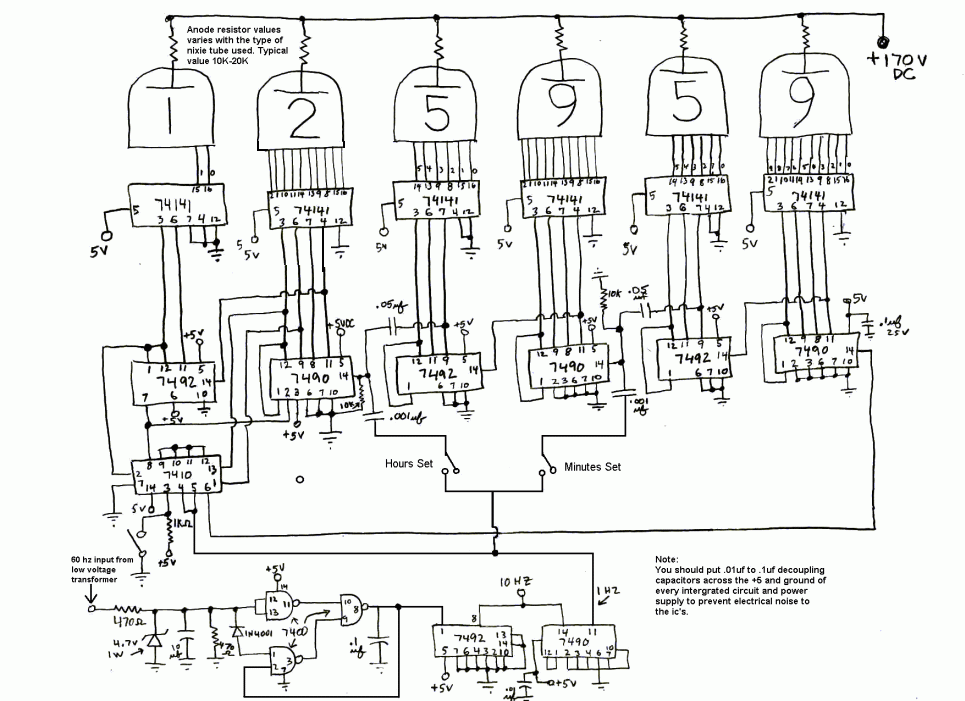 Это сделано для уменьшения количества микросхем и для облегчения трассировки платы. Впрочем, с моими стратегическими запасами микросхем К155ИД1 можно было бы сделать даже статическую хоть на 8 ламп. Лампы — ИН-12. Почему? Во-первых, у меня их штук двадцать. Во-вторых, мне так захотелось 🙂 В дальнем будущем планирую изготовить часы на лампах ИН-8-2, для которых ПП будет заказана на производстве. Короче, все серьезно. В сверхдальних планах часы на ИН-2. Четырех ламповые. Это сделано для уменьшения количества микросхем и для облегчения трассировки платы. Впрочем, с моими стратегическими запасами микросхем К155ИД1 можно было бы сделать даже статическую хоть на 8 ламп. Лампы — ИН-12. Почему? Во-первых, у меня их штук двадцать. Во-вторых, мне так захотелось 🙂 В дальнем будущем планирую изготовить часы на лампах ИН-8-2, для которых ПП будет заказана на производстве. Короче, все серьезно. В сверхдальних планах часы на ИН-2. Четырех ламповые.
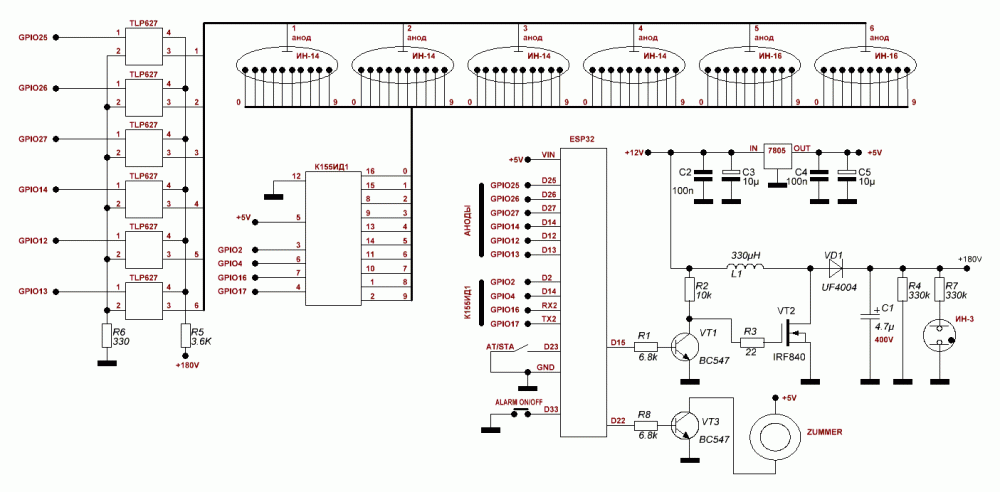
Ну-с, перейдем к делу. Сначала была разработана общая плата, на которой было все. И питание, и логика, и преобразователь. Но потом я нашел в нашей радиолаборатории при кружке деревянный корпус от конденсаторной батареи 64-года: Пришлось переработать плату, ибо она не влезала. Да и плату я загубил, забыв в УЗ ванне в спирте… Новая концепция: несколько отдельных плат, которые влезут в корпус. Схема осталась та же, просто конструкция стала блочной. Вот, например, уже изготовленный блок питания: Блок питания линейный. Изначально часы предполагались как четырехламповые, из-за чего было куплено 5 оптотранзисторов. Но ведь корпус такой огромный, тут только 6 ламп! Так как схема уже сделана, вытравлена и запаяна, то нужно придумать какой-нибудь костыль. Ну я и придумал. Сделал плату счетчик на советских микрухах — к155ие2 и ла3 и дешифратор ИД1. Плата получилась сложная, двух сторонняя. Начальная обработка корпуса состояла в сверлении отверстий под лампы. Для этого была проведена разметка. Сверление производилось в гараже, на станке сверлом 22,6. Окончательная обработка делалась напильником. Следующим этапом нужно было заделать все лишние отверстия в корпусе. Там были выводы конденсаторов. В отверстия вбивались обмазанные клеем китайские палочки, обрезались и сушились. Вот как это происходило: Потом наждачкой корпус был зачищен со всех сторон, останется загрунтовать его и покрасить. Нужно было как-то продумать крепление ламп. Нужно утопить их внутрь. Итак, я выпилил из тонкой фанеры пластинку длинной 24 см с прорезью под лампы по центру. Короче, обвел железное крепление самих ламп, которым они крепились в приборе. Чтобы сделать эту «деталь», пришлось вспомнить ручной лобзик, много лет пылящийся без дела, закупиться пилками для него (80р) и выпилить. Программирование. Весьма специфический этап. Тот момент, когда твой девайс оживает с залитием прошивки, которую прежде долго и муторно писал в Atmel Studio 6. С прошивкой было множество трудностей, часы никак не хотели работать так как надо. На индикаторы выводилась ерунда, динамическая индикация не работала. А все оказалось просто и банально — я неправильно работал с портами, совсем неправильно. Обнулял не правильно, из-за чего выводилось не то, что нужно. Так или иначе, с индикацией разобрался, но связи с микросхемой часов реального времени не было. Пришлось разбираться с шиной I2C и даташитом на DS3231… Доперев до того, что перед выводом на лампы, информацию о цифре нужно перекодировать в десятичный формат из двоично-десятичного, часы заработали. Сделал частичную шпаклевку для устранения дефектов корпуса в виде трещин, ямок и царапин. Высохнет — обработаем шкуркой, покрасим и можно устанавливать часы в корпус. Неужели. Покрасил корпус в черный цвет. Теперь, когда высохнет, надо будет смонтировать начинку в корпусе и все. 🙂 Когда корпус был готов, я стал разбираться с работой секунд. Они не работали. Был очень странный баг: секунды обнуляются, идут, но в определенный момент начинается случайный перебор всех значений счетчика и, как следствие, работать секунды адекватно не могут. Сначала я думал, что проблема аппаратная. Поэтому я напаял блокировочные конденсаторы на К155ИЕ2 и К155ЛА3. Поставил диод в цепь сброса, так как мне показалось, что выход И-НЕ, который сбрасывает счетчик при достижении 60 секунд будет конфликтовать с пином микроконтроллера. Сразу скажу сущность решения проблемы. У меня появилась мысль. А что, если ИЕ2 считает не по фронту, а по спаду? Начал спрашивать людей — все говорят, что логично по фронту. Стал читать интернет. Вычитал, что таки по спаду. Полный надежд побежал править код, поменял две строчки кода местами — заливаю прошивку и… оно работает! Всегда нужно бороться с предрассудками, истина — в технической документации и журналах «Радио». Все работает, можно устанавливать электронику в корпус и завершать проект. Внутри все прикручено мелкими саморезами. Плата логики и плата с DS3231 приклеены на термоклей. Для крепления трансформатора была вырезана полоска, чтобы закрепить тр-р. Сзади все просто. Вентиляционные отверстия, кнопки задания времени и разъем питания, совмещенный с держателем предохранителя. Итак. Проект часов на лампах ИН-12 завершен! Часы собраны, функционируют. Материалы
(54Kb) Печатные платы проекта в формате Sprint Layout 6 Домой — Статьи |
 В часы приходит 220 вольт через тумблер и предохранитель. Дальше ток идет через трансформатор ТС-4-1, вытащенный из убитого Океана-209, в диодный мост. Где-то вольт 18-20 попадает через кондеры на кренки, выдающие 5 и 12 вольт. 5 вольт нужно для питания логики, а 12В для питания преобразователя 12->180В выполненного на той же плате. Наружу 12 вольт не выходит. ШИМ для преобразователя берется с платы логики, с МК Atmega8. Так как блок линейный, то ему нужен радиатор. Силовой транзистор сидят на нем через изолирующие прокладки и вставки. Дело в том, что на корпусе у силового транзистора аж 180 вольт, а у кренок — земля.
В часы приходит 220 вольт через тумблер и предохранитель. Дальше ток идет через трансформатор ТС-4-1, вытащенный из убитого Океана-209, в диодный мост. Где-то вольт 18-20 попадает через кондеры на кренки, выдающие 5 и 12 вольт. 5 вольт нужно для питания логики, а 12В для питания преобразователя 12->180В выполненного на той же плате. Наружу 12 вольт не выходит. ШИМ для преобразователя берется с платы логики, с МК Atmega8. Так как блок линейный, то ему нужен радиатор. Силовой транзистор сидят на нем через изолирующие прокладки и вставки. Дело в том, что на корпусе у силового транзистора аж 180 вольт, а у кренок — земля. Заказал ее у китайцев, на производстве. Схема управляется импульсами: тактовый — прибавляет единицу, сброс — обнуляет, и питание. Вот что получилось:
Заказал ее у китайцев, на производстве. Схема управляется импульсами: тактовый — прибавляет единицу, сброс — обнуляет, и питание. Вот что получилось: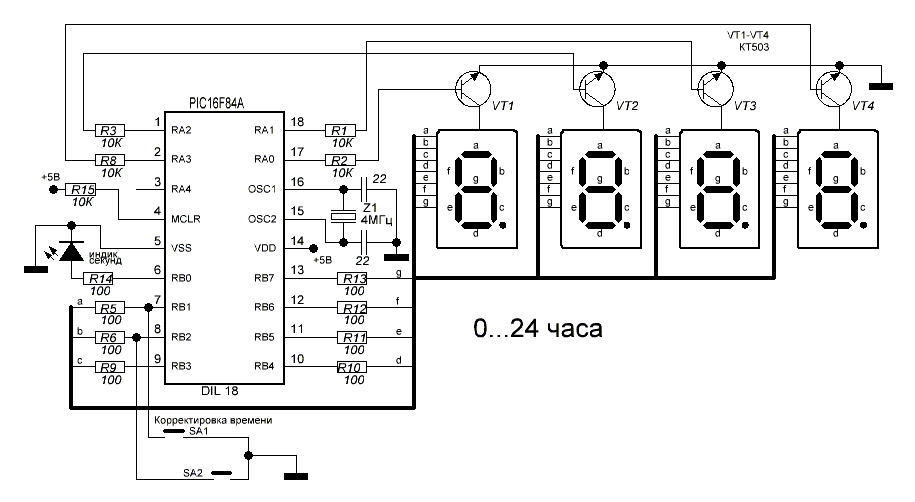 Как оказалось, пилить лобзиком я еще не разучился, что не может не радовать. Моей ручной сверлилкой были просверлены отверстия под болты для ламповых панелей. Закручены панели на их родные болты от прибора с гайками. Саму деталь можно легко приклеить изнутри. Но это позже. Вид:
Как оказалось, пилить лобзиком я еще не разучился, что не может не радовать. Моей ручной сверлилкой были просверлены отверстия под болты для ламповых панелей. Закручены панели на их родные болты от прибора с гайками. Саму деталь можно легко приклеить изнутри. Но это позже. Вид: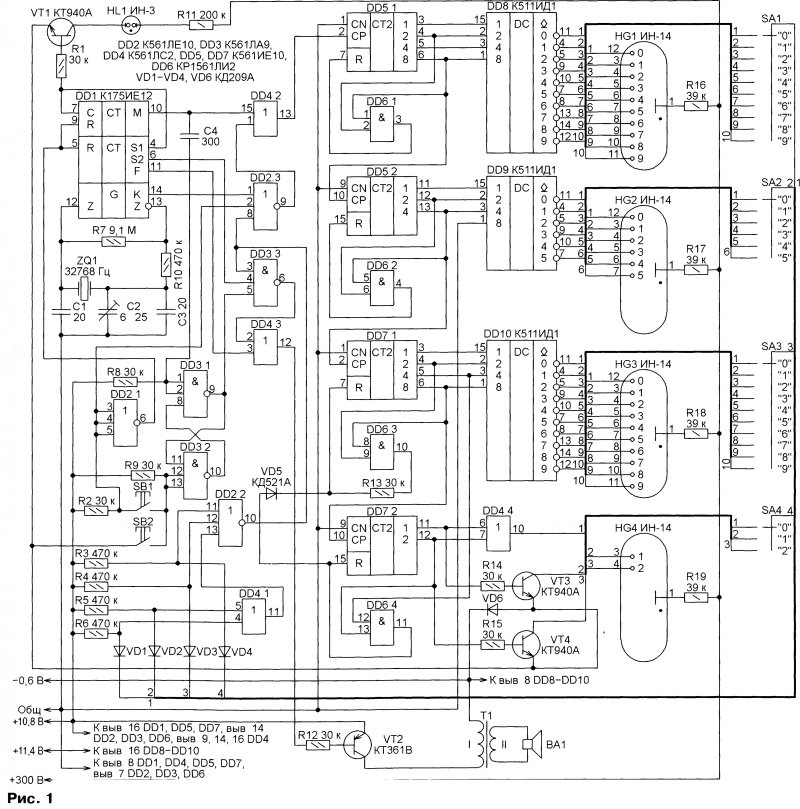 Какова была моя радость и восхищение, когда вместо 16 после 9-ти шла 10-ка! Причасные поймут. Итак, код отрабатывает. Часы идут.
Какова была моя радость и восхищение, когда вместо 16 после 9-ти шла 10-ка! Причасные поймут. Итак, код отрабатывает. Часы идут.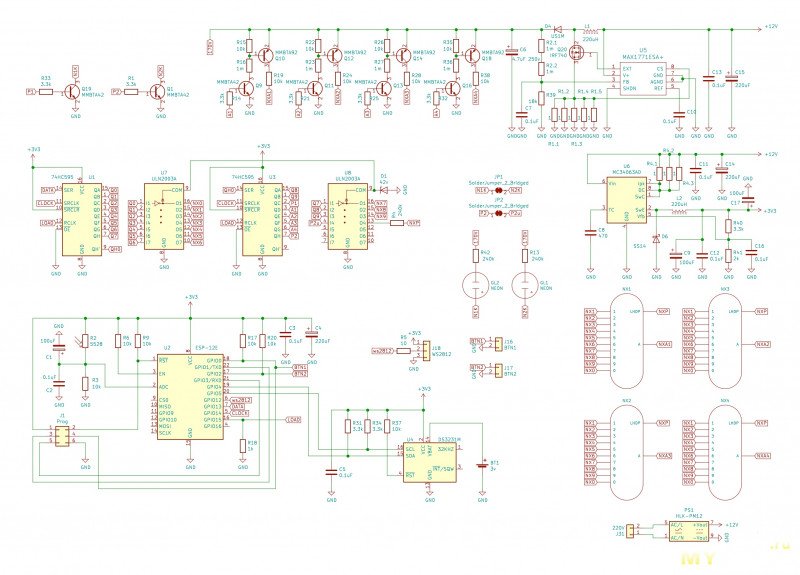 Напаял. Ничего не поменялось. Стал грешить на провода. Убираю провода — пересчета нет, но нет и счета. Итак, ошибка в программе.
Напаял. Ничего не поменялось. Стал грешить на провода. Убираю провода — пересчета нет, но нет и счета. Итак, ошибка в программе.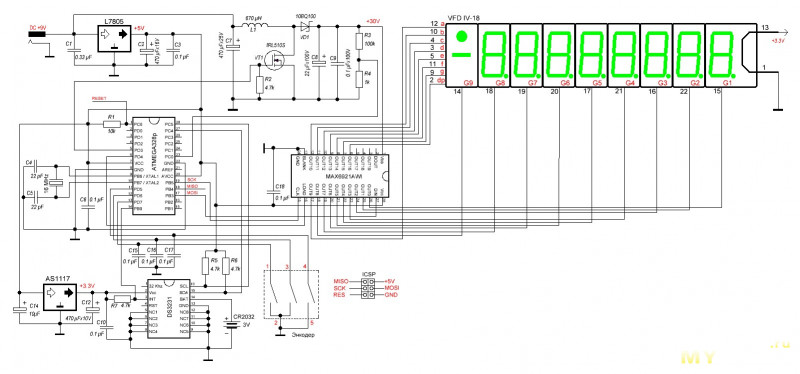
NiXIE: Симпатичные часы
Автор: wwakabobik
Лампа: ИН-1
Схема: есть
Плата:нет (монтажка)
Прошивка:есть
Исходник:есть
Описание: есть
Особенности: механический звонок.
Схема:
Файл схемы.
Наконец-то я доделал свой первый электротехнический проект: часы на газоразрядных индикаторах ИН-1 на базе Attiny x313 в около-стимпанковском стиле.
Всё началось с того, что случайно нагуглил по запросу “стимпанк” схожий девайс, а затем нашлась замечательная страничка на Radiokot. Насмотревшись красивых фотографий, подумалось мне: а почему бы не завести такое милое, рукодельческое хобби? Раньше мне никогда не приходилось заниматься электротехникой, и самое сложное, что я делал с паяльником — это чинил разъемы для наушников. Но это меня не остановило, хотя и внесло корректирующие требования: сложность схемы и пайки должна была быть простой, а с программой под девайс разберёмся. Всё-таки как-никак три года опыта с элитными и сложнейшими микроконтроллерами и встроенными системами. Поэтому был выбран обычный DIP-монтаж и схема на одной микросхеме: Attiny2313-20PU — найденная тут(упс,источник сдох).
Насмотревшись красивых фотографий, подумалось мне: а почему бы не завести такое милое, рукодельческое хобби? Раньше мне никогда не приходилось заниматься электротехникой, и самое сложное, что я делал с паяльником — это чинил разъемы для наушников. Но это меня не остановило, хотя и внесло корректирующие требования: сложность схемы и пайки должна была быть простой, а с программой под девайс разберёмся. Всё-таки как-никак три года опыта с элитными и сложнейшими микроконтроллерами и встроенными системами. Поэтому был выбран обычный DIP-монтаж и схема на одной микросхеме: Attiny2313-20PU — найденная тут(упс,источник сдох).
Первоначально я хотел реализовать проект 1 в 1, чтобы не усложнять себе жизнь, не имея опыта в сабже. Но быстро выяснилось, что, во-первых надо немного, но пошевелить мозгами, чтобы найти аналоги перечисленных в исходном случае микросхем. Во-вторых, почти сразу же меня стало не устраивать две вещи: 12-часовые часы, а не 24-часовые и отсутствие блинкующих разделителей. И сразу за двумя третье — если у нас есть будильник, то он должен быть не бездушным биппером, а полноценным звонком, механическим по мере возможностей.
И сразу за двумя третье — если у нас есть будильник, то он должен быть не бездушным биппером, а полноценным звонком, механическим по мере возможностей.
Определив техническое задание, были закуплены необходимые компоненты (большие и рабочие газоразрядные индикаторы пришлось поискать, и из желаемых ИН-16 и ИН-1 нашлись только последние; и то в количестве 7 штук, одна из которых оказалась битой). А так же разобрано три дисковых телефона, чтобы достать электромеханический, железный и звонкий звонок. А так же позаимствован у умершего роутера 12-вольтовый блок питания. Вся схема была перерисована мною в DipTrace, который оказался очень удачным как в плане структурной схемы, так и разводки платы. А так же заказана латунная фурнитура для ламп у хорошего мастера слесарных дел.
Всё необходимое можно найти в Чип и Дип’е за исключением газоразрядных индикаторов. Их можно найти под заказ в Кварце или на митинском радиорынке. Для монтажа мною была прикуплена паяльная станция и собственно макетная плата по грабительским расценкам. Впрочем, всё можно найти в разы дешевле, если задаться целью, а из Китая и вовсе в пару десятков вечнозелёных и убитых енотов. Я не буду приводить расписанную стоимость по каждой детали, отмечу только, что стоимость компонентов около 1т.р, ламп и платы — ещё 1,5т.р. и шкатулки (купленной на ярмарке в Измайловском кремле) с латунью — 1т.р. Итого 3-3,5 т.р. без паяльной станции и непредвиденных расходов.
Впрочем, всё можно найти в разы дешевле, если задаться целью, а из Китая и вовсе в пару десятков вечнозелёных и убитых енотов. Я не буду приводить расписанную стоимость по каждой детали, отмечу только, что стоимость компонентов около 1т.р, ламп и платы — ещё 1,5т.р. и шкатулки (купленной на ярмарке в Измайловском кремле) с латунью — 1т.р. Итого 3-3,5 т.р. без паяльной станции и непредвиденных расходов.
Непредвиденными оказались обстоятельства, что одна из оставшихся ламп треснула во время испытательного прогона, а две неоновых лампы, снятых с военной СТ-68 в своё время, сгорели из-за неправильно разведённой схемы — под них пришлось ставить 330кОм резисторы, при том, что дежурные индикаторы AM\PM, сделанные в Китае итак светят не столь ярко, как полагается неонкам. Совершенно неожиданным оказалось необходимость отказаться от варианта с лобзиком и напильником для работ по деревянной шкатулке, и приобрести сверло форестера, а так же удивительное ОТК СССР, которое пропускало все кривые и косые лампы с отклонениями по 1-2, а то и более миллиметров. Непредвиденным так же оказалось то, что электромеханический звонок не влез в шкатулку из-за чуть большего размера латунных втулок, из-за чего пришлось отложить вариант 12-вольтового питания с усиляющим конденсатором, и заменить его сложной системой звонка на основе виброзвонка и исходного купола звонка от телефона с возвратной пружиной. Впрочем, благодаря этому немного изменяя код, можно заставить будильник играть, к примеру, Чижика-Пыжика или симфониетту Янычека.
Непредвиденным так же оказалось то, что электромеханический звонок не влез в шкатулку из-за чуть большего размера латунных втулок, из-за чего пришлось отложить вариант 12-вольтового питания с усиляющим конденсатором, и заменить его сложной системой звонка на основе виброзвонка и исходного купола звонка от телефона с возвратной пружиной. Впрочем, благодаря этому немного изменяя код, можно заставить будильник играть, к примеру, Чижика-Пыжика или симфониетту Янычека.
Во время замены треснувшей ИН-1 оказалось, что не все ИН-1 одинаковы, и у приобретённых в Митино ламп оказался шаг сетки меньше, чем у приобретённых в Кварце, что визуально бросалось в глаза. Поэтому пришлось лампы перезаказать ещё раз в Кварце. Так же оказалось, что код, позволяющий выводить дату, физически не помещается в 2Кб памяти. Поэтому было найдено решение использовать ту же разводку, но на базе Attiny4313 с 4Кб памяти. Код так же пришлось сильно оптимизировать и трюкачествовать, хотя всё ещё на си, допускающем и большие оптимизации, не говоря о компактности ассемблера. Но, проще взять схожую микросхему, чем биться за каждый байт. Впрочем, все усложнения кода привели к тому, что все мои действия не влезли в исходный фрейм 1мс и происходил overrun, выливающийся в подвисание кнопок и зависании будильника. Поэтому фрейм был сменён на 2мс, что никак визуально не повлияло на восприятие часов. Тем не менее, несмотря на все злоключения, AVR Studio мне очень сильно помогла и своим встроенным дебаггером (конечно с Dragon, наверное было бы удобнее, но, увы, на один раз я не готов покупать отладочную плату) и своими просто замечательными оптимизациями.
Но, проще взять схожую микросхему, чем биться за каждый байт. Впрочем, все усложнения кода привели к тому, что все мои действия не влезли в исходный фрейм 1мс и происходил overrun, выливающийся в подвисание кнопок и зависании будильника. Поэтому фрейм был сменён на 2мс, что никак визуально не повлияло на восприятие часов. Тем не менее, несмотря на все злоключения, AVR Studio мне очень сильно помогла и своим встроенным дебаггером (конечно с Dragon, наверное было бы удобнее, но, увы, на один раз я не готов покупать отладочную плату) и своими просто замечательными оптимизациями.
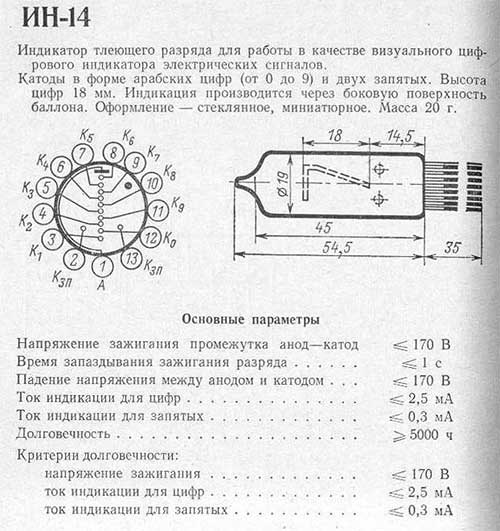
Отдельно хочу сказать про техническую сторону дела. Монтаж я производил исключительно проводами\ножками, т.к. протравливать плату — ещё дополнительные расходы на реагенты, и для разового проекта неразумны. Лампы достаточно капризны в плане пайки и монтажа, поэтому к ним надо проявлять особо бережное отношение.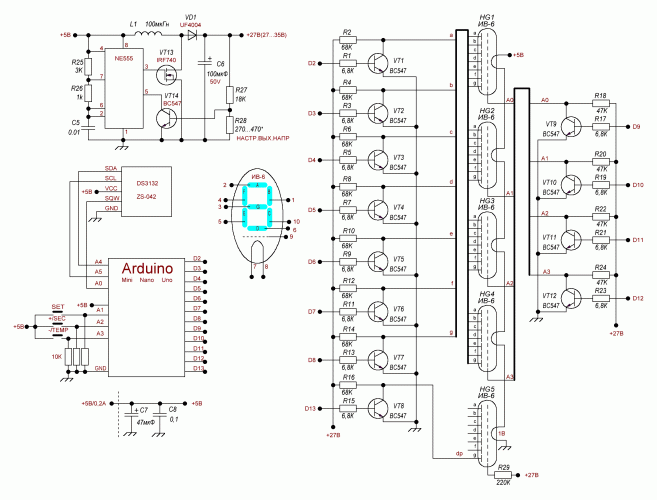 Электрическая же схема очень хорошая, во-первых, не раз получая КЗ из-за выгоревших, а затем и треснувшей лампы, из-за замкнувшей ножки схемы и микросхемы отлично себя чувствовали несмотря ни на что. Отчасти благодаря грамотным транзисторным ключам, и отчасти силовой электроники. Поэтому особое внимание следует обратить на стабилизационный контур, а так же мощность элементов. Хотя в оригинале товарищ рекомендует использовать 250В конденсатор и 200мкГн элемент индуктивности, тем не менее, в моём случае 400В и 250мкГн работают только лучше. И отдельно хочу сказать про потребление и надёжность. Во-первых часы работают в импульсном режиме со скважностью 6. Поэтому и потребление, и выгорание ламп, и тепловыделение почти незаметное. Сам не измерял, но все элементы комнатной температуры, так что можно верить в то, что оно не больше 2-3Вт.
Электрическая же схема очень хорошая, во-первых, не раз получая КЗ из-за выгоревших, а затем и треснувшей лампы, из-за замкнувшей ножки схемы и микросхемы отлично себя чувствовали несмотря ни на что. Отчасти благодаря грамотным транзисторным ключам, и отчасти силовой электроники. Поэтому особое внимание следует обратить на стабилизационный контур, а так же мощность элементов. Хотя в оригинале товарищ рекомендует использовать 250В конденсатор и 200мкГн элемент индуктивности, тем не менее, в моём случае 400В и 250мкГн работают только лучше. И отдельно хочу сказать про потребление и надёжность. Во-первых часы работают в импульсном режиме со скважностью 6. Поэтому и потребление, и выгорание ламп, и тепловыделение почти незаметное. Сам не измерял, но все элементы комнатной температуры, так что можно верить в то, что оно не больше 2-3Вт.
И, наконец, исходный код, бинарники и схему можно найти тут для 2313 и 4313.
Зеркало 2313 и 4313.
Первоисточник.
15. Часы — Джон Книн: Микроконтроллеры
 w3.org/1999/xhtml» cellspacing=»0″>
w3.org/1999/xhtml» cellspacing=»0″>- Введение и важность синхронизации действий.
- Базовые схемы часов
- Такты микроконтроллера ARM.
Материал для конкретного устройства
- NXP LPC1768 Выбор часов. Устройство, используемое в MBED. LPC1768 Пример: тактовая частота 48 МГц для USB
- STM32F107 Выбор часов. Устройство, используемое в оценочной плате Keil MCBSTM32C.
- STM32F407 Выбор часов. Устройство, используемое в Discovery Board
Знакомство с часами и временем.
Работа микроконтроллера синхронизируется с системными часами. На уровне ЦП выборка, декодирование и выполнение инструкций микроконтроллера происходят во время тактового импульса микроконтроллера. На системном уровне все действия/действия микроконтроллера синхронизируются с синхронизирующими сигналами, которые устанавливаются на фиксированное число, кратное основному тактовому периоду.
На этой странице будут рассмотрены часы микроконтроллера. Таймеры рассматриваются на отдельных страницах.
- Важность синхронизации действий (неэлектронные примеры). Время — фундаментальное свойство, которое измеряется часами. На рабочем месте жизнь людей определяется временем начала, перерыва и окончания. Затем время общественного транспорта диктует, когда они должны выходить из дома и когда они прибывают домой, что, в свою очередь, диктует их свободное время. Если они должны встретиться с кем-то, то они оба должны договориться о каком-то взаимоприемлемом времени.
Все эти действия основаны на приборах для измерения времени, т.е. синхронизируемых часах. События, приведшие к фатальному полету знаменитой летчицы Амелии Эрхарт в 1937 году из Лаэ, Новая Гвинея, на остров Хауленд в Тихом океане, были омрачены недопониманием и путаницей в отношении времени контакта. Поскольку она путешествовала с запада на восток через множество часовых поясов, возникла значительная путаница в отношении того, было ли время местным временем в ее пункте отправления, ее пункте прибытия, или это было местное время того, для кого она общалась.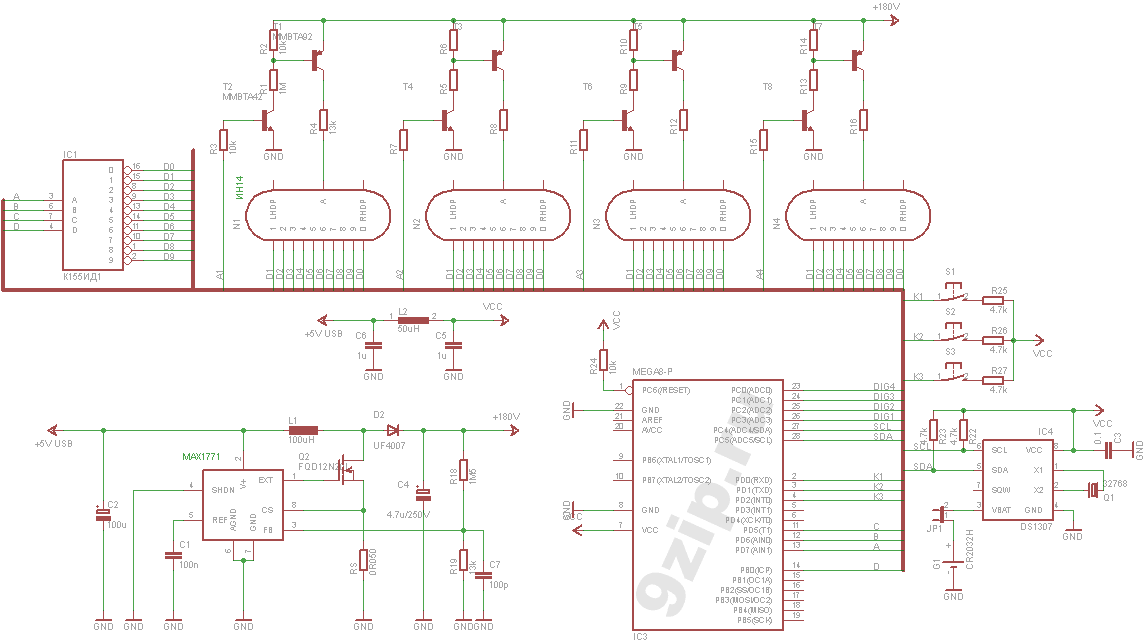 Кроме того, поскольку часть ее поддержки базировалась на Гавайях, которые находились по другую сторону от международной линии перемены дат, возникла путаница в отношении того, какой сегодня день.
Кроме того, поскольку часть ее поддержки базировалась на Гавайях, которые находились по другую сторону от международной линии перемены дат, возникла путаница в отношении того, какой сегодня день.
Примечания
- Ссылка «Амалия Эрхарт: Тайна раскрыта» Эген М. Лонг и Мари К. Лонг. Саймон и Шустер в мягкой обложке © 1999
- Все контакты должны быть организованы с помощью телеграфа и иметь вид «Свяжусь с вами в 09:00 по любому поводу». Время оборота телеграфа было настолько долгим, что подтверждение было невозможно. Кроме того, когда была предпринята попытка установить радиосвязь, и оборудование, и прием были настолько плохими, что легко было предположить, что радио не работает в назначенное время, и просто нажать, предполагая, что все в порядке.
Маршрут рокового полета Амелии Эрхарт.
Поскольку расследования после гибели людей, особенно в получивших широкую огласку авариях, приводят к улучшениям после этого и других подобных событий, вся авиация теперь работает по всемирному времени или среднему времени по Гринвичу.
Для определения своего положения кораблям необходим точный хронометр. Во многих портах была шаровая башня, соединенная с обсерваторией. Ровно в 13:00 каждый день шар поднимали на вершину башни и опускали, позволяя штурманам кораблей синхронизировать свои хронометры.
Ball Towers в Гринвиче, Лондон и Уильямстаун, Мельбурн.
В предыдущем разделе была показана необходимость измерения времени. Системы микроконтроллеров включают в себя схемы, которые генерируют повторяющийся сигнал, известный как часы. Простые системы будут основаны на RC-цепи, а системы, требующие большей точности, будут включать кристалл. В то время как микроконтроллер может включать часть необходимой схемы внутри из-за производственных ограничений, конденсаторы и кристаллы будут внешними.
Простая схема часов на кристалле.
В приведенной выше схеме первые два логических элемента образуют неинвертирующий усилитель.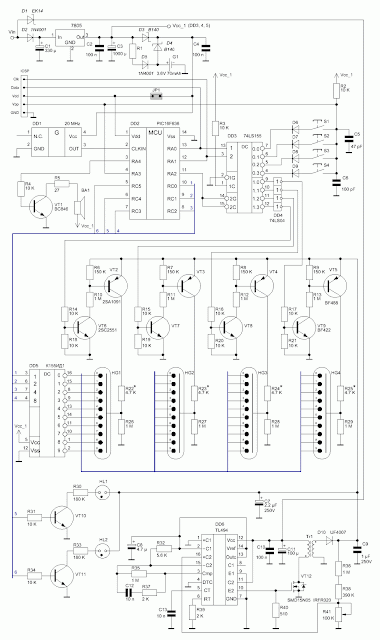 Любой шум на входе будет усиливаться, заставляя схему колебаться. Поскольку кристалл имеет очень острую частотную характеристику, конструкция колеблется на частоте кристалла. На входе усилителя сигнал будет синусоидой плохого качества, тогда как на выходе усилителя сигнал может быть не очень качественным прямоугольным сигналом. Третий логический вентиль должен обеспечивать идеальную прямоугольную волну на частоте кварца.
Любой шум на входе будет усиливаться, заставляя схему колебаться. Поскольку кристалл имеет очень острую частотную характеристику, конструкция колеблется на частоте кристалла. На входе усилителя сигнал будет синусоидой плохого качества, тогда как на выходе усилителя сигнал может быть не очень качественным прямоугольным сигналом. Третий логический вентиль должен обеспечивать идеальную прямоугольную волну на частоте кварца.
Часы ARM. STM32F105xx, STM32F107xx Устройства линии связи
Как показано, микроконтроллер ARM имеет четыре источника синхронизации, два внутренних и два внешних.
| Часы | NXP LPC1768 | СТМ32Ф107 | СТМ32Ф407 | Комментарии |
|---|---|---|---|---|
| используется в плате MBED | используется в Keil MCBSTM32C | используется в Discovery Board | ||
| Внутренний RC-генератор | 4 МГц | часы по умолчанию после сброса.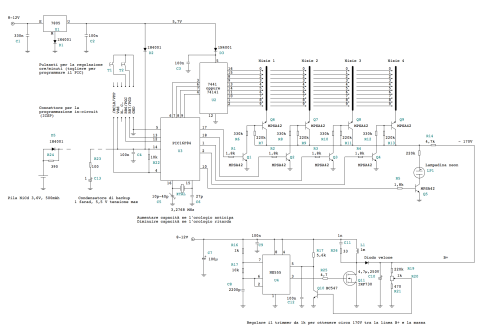 Не рекомендуется для точных частот. Не рекомендуется для точных частот. | ||
| Высокоскоростные внутренние часы (HSI). | 8 МГц | 16 МГц | часы по умолчанию после сброса | |
| Генератор часов реального времени | 1 Гц для часов реального времени; 32 кГц для WDT | используется сторожевым таймером | ||
| Низкоскоростные внутренние часы (LSI) | 40 кГц | 32 кГц | используется сторожевым таймером | |
| Часы-генератор | 12 МГц в комплекте | Набор внешних кристаллов. 1-25 для NXP LPC1768 | ||
| Высокоскоростные внешние часы (HSE) | 25 МГц в комплекте | 8 МГц поставляется | Набор внешних кристаллов. 3-25 для STM32F107 и 4-26 для STM32F407 | |
| Низкоскоростные внешние часы (LSE) | 32,768 кГц | Кристалл не входит в комплект поставки | Используется для генерации внутренних RTC (Часов Реального Времени). С делителем на 14 ступеней выдается 1-секундный сигнал. |
Часы LSI имеют номинальную частоту (40 кГц для STM32F107), но могут варьироваться от 30 до 60 кГц из-за производственных допусков. HSI был отрегулирован лазером, поэтому он намного ближе к своему номинальному значению (8 МГц для STM32F107). Если для внешних часов используются кристаллы, как показано на плате Keil ARM ниже, эти часы будут очень точными (1 часть из 10 9).0204 7 ).
Расположение кристаллов на плате Keil ARM MCBSTM32C.
ARM также имеет выход тактового сигнала MCO (выход тактового сигнала микроконтроллера), который можно запрограммировать для вывода одного из многих внутренних часов ARM.
LSE и MCO — это альтернативные функции, которые используют общие контакты ввода-вывода микроконтроллера.
Создано John Kneen RMIT University суббота, 29 июня 2013 г. Обновлено пятница, 8 ноября 2013 г.
Оборудование — часы и восстановление данных
Аппаратное обеспечение – часы и восстановление данных Джон Харрингтон Комментарии Восстановление часов и данных — важная функция физического уровня современных коммутаторов и маршрутизаторов. Возможно, вам не по душе копаться в электронных компонентах маршрутизатора, но восстановление тактовой частоты является фундаментальным строительным блоком для других функций сетевого оборудования. Например, для эффективного преобразования последовательных данных в параллельные требуются надежные часы и восстановление данных (CDR). Трудно понять последовательное преобразование в параллельное или преобразование сигнала, не изучив предварительно CDR.
С точки зрения MAC-уровня передатчика скорость передачи 10GBase-R составляет 10 Гбит/с. (10Gbase-R здесь относится к SR, LRM, LR, ER и т. д.) Однако на физической линии скорость передачи составляет 10,3125 Гбит/с. Канал 10Gbase-R использует кодирование 64b/66b, которое делит данные уровня 2, полученные от уровня MAC, на 64-битные блоки и вставляет дополнительные 2 бита заголовка перед каждым блоком. Эти накладные расходы не расходуют доступную полосу пропускания. Вместо этого передатчик использует «разгон» и передает закодированный битовый поток, используя тактовую частоту 10,3125 кГц. Эти дополнительные биты удаляются принимающими цепями физического уровня, поэтому линейное кодирование и разгон не видны на уровне MAC. Кроличья нора в кодировке 64b/66b гораздо глубже, но пока хорошо знать, что существует кодировка 64b/66b.
В цифровом приемнике 10GBase-R полученный сигнал интерпретируется путем выборки формы сигнала прямо в середине ожидаемого битового периода. Чтобы выровнять эту выборку и правильно декодировать полученный сигнал как единицу или ноль, вам необходимо иметь часы приема, которые работают точно с той же частотой и фазой, что и часы передатчика. К сожалению, вы не можете просто установить тот же генератор 10,3125 кГц в ресивер и покончить с этим. Нет… это было бы слишком просто….
К сожалению, вы не можете просто установить тот же генератор 10,3125 кГц в ресивер и покончить с этим. Нет… это было бы слишком просто….
Загвоздка в том, что ни один осциллятор не совершенен. Стандарт 802.3 10G обеспечивает точность часов в пределах +/- 0,01 процента. Поэтому вполне вероятно, что часы передачи и приема будут рассинхронизированы. Это называется тактовым или частотным дрейфом. Несоответствие часов приведет к потере битов из-за проскальзывания или ложной интерпретации битов, когда приемник производит выборку в неправильной точке формы волны, путая «единицу» с «нулем» или наоборот.
На приведенной ниже диаграмме показано, что тактовый сигнал Rx (красный) работает быстрее, чем тактовый сигнал Tx и сигнал данных. Хотя фазы первых переходов 0-1 совпадают по фазе между двумя сигналами, в течение нескольких тактов они полностью противофазны. Да, это преувеличение, но это реальная проблема.
Даже если часы Tx и Rx работают на одной и той же частоте, их все равно необходимо выровнять, чтобы их фазы совпадали.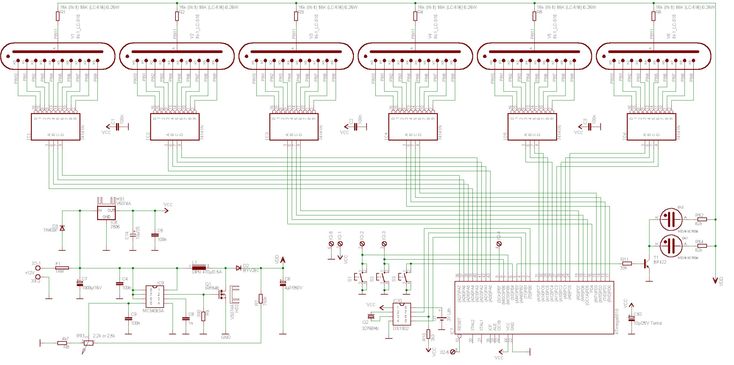 Вы можете увидеть диаграмму ниже, показывающую, что часы работают на одной частоте, но не совпадают по фазе друг с другом, что снова приводит к неправильной выборке принятого сигнала.
Вы можете увидеть диаграмму ниже, показывающую, что часы работают на одной частоте, но не совпадают по фазе друг с другом, что снова приводит к неправильной выборке принятого сигнала.
«У него….. захват нас! – Он увлекает меня. Проклятье. Мустанг, это пугало вокруг меня»
Вместо того, чтобы пытаться угадать точную частоту и фазу передатчика, почему бы не определить их по входящему сигналу? Это восстановление часов и данных (CDR). Функция CDR, встроенная в приемник, имеет собственный генератор и использует умный компонент, называемый контуром фазовой автоподстройки частоты (PLL), для калибровки генератора и его согласования с фазой и частотой входящего сигнала. В PLL есть «фазовый компаратор», который сравнивает фазу принимаемого сигнала с фазой гетеродина. Разница между двумя сигналами создает выходной сигнал, который приводит в действие генератор, управляемый напряжением (VCO). Этот вход управления VCO модифицирует генератор, чтобы он лучше соответствовал входящему сигналу, и подает локальный сигнал обратно в фазовый компаратор, чтобы снова начать процесс.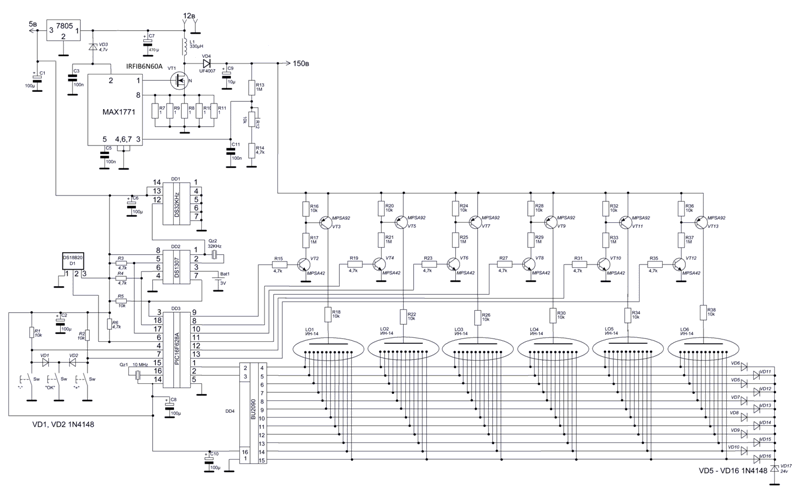
Я знаю, я теряю тебя, но мы закончили с сумасшедшей электроникой. Цепь обратной связи в конечном итоге синхронизирует гетеродин с фазой и частотой входящего сигнала. Вот и все. Как только ваши часы PLL «заблокированы», приемник может правильно рассчитать время выборки для восстановления переданных данных. Тем не менее, часы передатчика могут изменять свою частоту в зависимости от времени и температуры, поэтому PLL должен поддерживать эту синхронизацию, постоянно контролируя принимаемый сигнал.
Я замахнулся на важный элемент часов и восстановления данных — переходы между уровнями. Переход уровня — это изменение от единичного значения к нулевому значению и от нуля к единице. В приведенных выше примерах поток битов представлял собой идеалистическую прямоугольную волну 1-0-1-0-1-0. В реальном мире, если в принятом потоке данных нет переходов уровней (например, поток всех единиц или всех нулей), тогда PLL теряет способность определять тактовую частоту передатчика. Это называется «потеря блокировки». «Длина цикла» PLL — это мера того, как долго он может выдерживать отсутствие переходов, прежде чем потеряет синхронизацию.
«Длина цикла» PLL — это мера того, как долго он может выдерживать отсутствие переходов, прежде чем потеряет синхронизацию.
: Передаваемый сигнал необходимо каким-то образом изменить, чтобы помочь приемнику восстановить и сохранить синхронизацию часов. В 10Base-T Ethernet использовалось манчестерское кодирование, которое гарантировало переход уровня в течение каждого битового времени. К сожалению, он сделал это, удвоив тактовую частоту линии, используя два тактовых импульса для каждого передаваемого бита данных. Это 100% накладные расходы на линейное кодирование.
В 10GBase-R эта проблема решается линейным кодированием 64b/66b. Два служебных бита (или преамбулы) могут иметь только значения 0-1 или 1-0. Преамбула 0-1 означает, что следующий 64-битный блок содержит чистые данные, а преамбула 1-0 означает, что блок содержит некоторую управляющую информацию. Мы рассмотрим сигнализацию позже, но учтите, что шаблоны преамбулы 0-1 и 1-0 «гарантируют» переход уровня через каждые 64 бита. Таким образом, мы можем сказать, что 10GBase-R имеет максимальную 64-битную «длину выполнения». Накладные расходы на линейное кодирование составляют 3,125%, что более приемлемо, чем накладные расходы на манчестерское кодирование.
Таким образом, мы можем сказать, что 10GBase-R имеет максимальную 64-битную «длину выполнения». Накладные расходы на линейное кодирование составляют 3,125%, что более приемлемо, чем накладные расходы на манчестерское кодирование.
Кроме того, 64-битный блок «зашифрован» для обеспечения дальнейших переходов между уровнями. Это делается не только для восстановления тактовой частоты и делает очень вероятным то, что у вас будут переходы уровней в вашем потоке данных, поэтому хорошо иметь принудительные переходы из 2-битных преамбул.
Когда полученный последовательный битовый поток 10GBase-R должен быть передан на уровень MAC, он должен быть преобразован для передачи на уровень MAC с использованием 32-битного параллельного интерфейса интерфейса XGMII. Преобразователь Serial-in/Parallel-out (SIPO) должен выполнять здесь CDR. Восстановленные биты данных синхронизируются в 32-битном регистре с линейной скоростью. Восстановленные часы делятся на 32 и используются для синхронизации 32-битных слов на шине с уменьшенной скоростью передачи данных.
Иногда требуется выполнить CDR, даже если вы планируете повторно передать полученные последовательные данные как другой поток последовательных данных. Это форма переобучения, известная как повторное определение времени. Сигналы, передаваемые в канале, будут ослабляться (уменьшаться в размере) и искажаться (изменяться в форме) по мере их распространения, и, как обсуждалось в предыдущем посте, печатная плата представляет собой враждебную среду. Восстанавливая часы и данные, а затем регенерируя импульсы, вы можете продлить срок службы сигнала.
Наконец, следует отметить, что механизм CDR обычно имеет скрытый компаньон, называемый эквалайзером или эквалайзером. Эквалайзер — это еще одна часть обработки сигнала, усиливающая высокочастотные компоненты принятого сигнала, очищающая его, чтобы схема CDR могла иметь шанс определить переходы уровней. Скоро мы рассмотрим схемы эквалайзера.
Если у вас есть какие-либо вопросы, исправления или общие отзывы, дайте мне знать в комментариях.
аппаратное обеспечение
10G, 64b/66b, CDR, EQ, PLL, синхронизация, VCO
Цепь гена циркадных часов контролирует белковые и фосфопротеиновые ритмы у Arabidopsis thaliana
, Холлидей К.Дж., Миллар А.Дж. Цепь часового гена у арабидопсиса включает репрессилятор с дополнительными петлями обратной связи. Мол. Сист. биол. 2012; 8:574. [Бесплатная статья PMC] [PubMed] [Google Scholar]
2. Fogelmark K., Troein C. Переосмысление активации транскрипции в циркадных часах арабидопсиса. PLoS-компьютер. биол. 2014;10 [Бесплатная статья PMC] [PubMed] [Google Scholar]
3. van Ooijen G., Millar A.J. Нетранскрипционные осцилляторы в циркадном хронометраже. Тенденции биохим. науч. 2012; 37: 484–492. [PubMed] [Google Scholar]
4. Daniel X., Sugano S., Tobin E.M. Фосфорилирование CK2 CCA1 необходимо для его функции циркадного осциллятора у арабидопсиса. проц. Натл. акад. науч. США 2004; 101:3292–3297. [Бесплатная статья PMC] [PubMed] [Google Scholar]
5. Нишиваки Т., Сатоми Ю., Накадзима М., Ли С., Киёхара Р., Кагеяма Х., Китаяма Ю., Темамото М., Ямагути А. ., Hijikata A., Go M., Iwasaki H., Takao T., Kondo T. Роль фосфорилирования KaiC в системе циркадных часов Synechococcus elongatus PCC 7942. Proc. Натл. акад. науч. США 2004; 101:13927–13932. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Нишиваки Т., Сатоми Ю., Накадзима М., Ли С., Киёхара Р., Кагеяма Х., Китаяма Ю., Темамото М., Ямагути А. ., Hijikata A., Go M., Iwasaki H., Takao T., Kondo T. Роль фосфорилирования KaiC в системе циркадных часов Synechococcus elongatus PCC 7942. Proc. Натл. акад. науч. США 2004; 101:13927–13932. [Бесплатная статья PMC] [PubMed] [Google Scholar]
6. Майер Б., Вендт С., Ванселов Дж. Т., Уоллах Т., Райшл С., Эмке С., Шлоссер А., Крамер А. Крупномасштабный Функциональный скрининг РНКи показывает роль CK2 в циркадных часах млекопитающих. Гены Дев. 2009 г.;23:708–718. [Бесплатная статья PMC] [PubMed] [Google Scholar]
7. Yang Y., Cheng P., He Q., Wang L., Liu Y. Фосфорилирование частотного белка казеинкиназой II необходимо для функции Циркадные часы нейроспоры. Мол. Клетка. биол. 2003; 23:6221–6228. [Бесплатная статья PMC] [PubMed] [Google Scholar]
8. Кусакина Дж., Додд А.Н. Фосфорилирование в циркадной системе растений. Тенденции Растениевод. 2012; 17: 575–583. [PubMed] [Google Scholar]
9. Чоудхари М.К., Номура Ю., Ван Л., Накагами Х., Сомерс Д.Э. Количественный циркадный фосфопротеомный анализ арабидопсиса выявляет обширный часовой контроль ключевых компонентов физиологических, метаболических и сигнальных путей. Мол. Клетка. Протеомика. 2015;14:2243–2260. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Чоудхари М.К., Номура Ю., Ван Л., Накагами Х., Сомерс Д.Э. Количественный циркадный фосфопротеомный анализ арабидопсиса выявляет обширный часовой контроль ключевых компонентов физиологических, метаболических и сигнальных путей. Мол. Клетка. Протеомика. 2015;14:2243–2260. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Лелуп Дж. К., Голдбитер А. На пути к подробной вычислительной модели циркадных часов млекопитающих. проц. Натл. акад. науч. США 2003; 100:7051–7056. [Бесплатная статья PMC] [PubMed] [Google Scholar]
11. Бейкер К.Л., Лорос Дж.Дж., Данлэп Дж.К. Циркадные часы Neurospora crassa. ФЭМС микробиол. 2012; 36:95–110. [Бесплатная статья PMC] [PubMed] [Google Scholar]
12. Takahashi J.S. Транскрипционная архитектура циркадных часов млекопитающих. Нац. Преподобный Жене. 2017;18:164–179. [Бесплатная статья PMC] [PubMed] [Google Scholar]
13. Mauvoisin D., Wang J., Jouffe C., Martin E., Atger F., Waridel P., Quadroni M., Gachon F., Naef F. Зависимые от циркадных часов и -независимые ритмические протеомы реализуют различные суточные функции в печени мышей.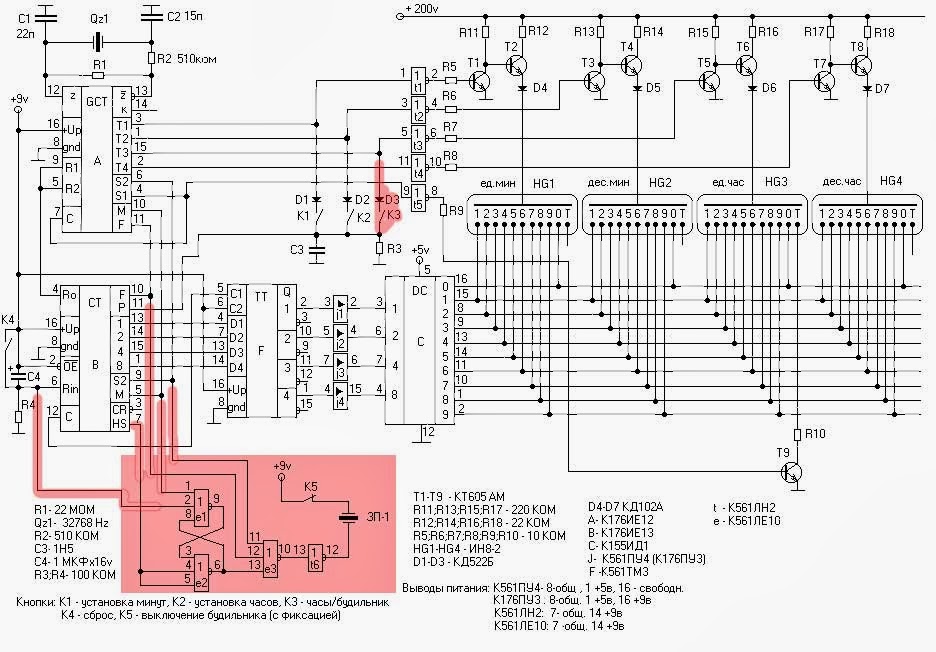 проц. Натл. акад. науч. США 2014; 111:167–172. [Бесплатная статья PMC] [PubMed] [Google Scholar]
проц. Натл. акад. науч. США 2014; 111:167–172. [Бесплатная статья PMC] [PubMed] [Google Scholar]
14. Люк С., Терли К., Табен П.Ф., Вестермарк П.О. Ритмическая деградация объясняет и объединяет циркадные данные транскриптома и протеома. Представитель ячейки 2014; 9: 741–751. [PubMed] [Google Scholar]
15. Чоудхари М.К., Номура Ю., Ши Х., Накагами Х., Сомерс Д.Э. Циркадное профилирование протеома арабидопсиса с использованием 2D-DIGE. Фронт. Растениевод. 2016;7:1007–1014. [Бесплатная статья PMC] [PubMed] [Google Scholar]
16. Накадзима М., Имаи К., Ито Х., Нишиваки Т., Мураяма Ю., Ивасаки Х., Ояма Т., Кондо Т. Восстановление циркадного ритма колебания фосфорилирования KaiC цианобактерий in vitro. Наука. 2005; 308: 414–415. [PubMed] [Академия Google]
17. О’Нил Дж.С., ван Ойен Г., Диксон Л.Е., Троейн С., Кореллоу Ф., Буге Ф.Ю., Редди А.Б., Миллар А.Дж. Циркадные ритмы сохраняются без транскрипции у эукариот. Природа. 2011; 469: 554–558. [Бесплатная статья PMC] [PubMed] [Google Scholar]
18. O’Neill J.S., Reddy A.B. Циркадные часы в эритроцитах человека. Природа. 2011; 469: 498–503. [Бесплатная статья PMC] [PubMed] [Google Scholar]
O’Neill J.S., Reddy A.B. Циркадные часы в эритроцитах человека. Природа. 2011; 469: 498–503. [Бесплатная статья PMC] [PubMed] [Google Scholar]
19. Эдгар Р.С., Грин Э.В., Чжао Ю., Ван Оойен Г., Олмедо М., Цинь Х., Сюй Ю., Пан М., Валекунья Великобритания, Фини К.А., Мэйвуд Э.С., Гастингс М.Х., Балига Н.С., Мерроу М., Миллар А.Дж. и др. Пероксиредоксины являются консервативными маркерами циркадианных ритмов. Природа. 2012;485:459–464. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Фини К.А., Хансен Л.Л., Путкер М., Оливарес-Яньес К., Дэй Дж., Идес Л.Дж., Ларрондо Л.Ф., Хойл Н.П., О’Нил Дж.С., van Ooijen G. Ежедневные потоки магния регулируют клеточный хронометраж и энергетический баланс. Природа. 2016; 532: 375–379. [Бесплатная статья PMC] [PubMed] [Google Scholar]
21. Henslee EA, Crosby P., Kitcatt S.J., Parry JSW, Bernardini A., Abdallat R.G., Braun G., Fatoyinbo H.O., Harrison E.J., Edgar R.S., Hoettges К.Ф., Редди А.Б., Джабр Р.И., фон Шанц М., О’Нил Дж. С. и др. Ритмичный транспорт калия регулирует циркадные часы в эритроцитах человека. Нац. коммун. 2017;8:1978. [Бесплатная статья PMC] [PubMed] [Google Scholar]
С. и др. Ритмичный транспорт калия регулирует циркадные часы в эритроцитах человека. Нац. коммун. 2017;8:1978. [Бесплатная статья PMC] [PubMed] [Google Scholar]
22. Редди А.Б., Карп Н.А., Мэйвуд Э.С., Сейдж Э.А., Дири М., О’Нил Дж.С., Вонг Г.К., Чешам Дж., Оделл М., Лилли К.С., Кириаку С.П., Гастингс М.Х. Циркадная оркестровка печеночного протеома. Курс. биол. 2006; 16:1107–1115. [PubMed] [Google Scholar]
23. Роблес М.С., Кокс Дж., Манн М. Количественная протеомика in vivo выявляет ключевой вклад посттранскрипционных механизмов в циркадную регуляцию метаболизма печени. Генетика PLoS. 2014;10 [Бесплатная статья PMC] [PubMed] [Google Scholar]
24. Роблес М.С., Хамфри С.Дж., Манн М. Фосфорилирование является центральным механизмом циркадного контроля метаболизма и физиологии. Клеточный метаб. 2017;25:118–127. [PubMed] [Google Scholar]
25. Уриг Р.Г., Эчеварриа-Зоменьо С., Шлапфер П., Гроссманн Дж., Рощицки Б., Кербер Н., Фиорани Ф., Груиссем В. Суточная динамика протеома розетки арабидопсиса и фосфопротеом. Окружающая среда растительной клетки. 2021; 44: 821–841. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Окружающая среда растительной клетки. 2021; 44: 821–841. [Бесплатная статья PMC] [PubMed] [Google Scholar]
26. Уриг Р.Г., Шлепфер П., Рошицки Б., Хирш-Хоффманн М., Груиссем В. Суточные изменения согласованного фосфорилирования и ацетилирования растительных белков в органах и проростках арабидопсиса . Завод Ж. 2019;99:176–194. [PubMed] [Google Scholar]
27. Wang Z.Y., Tobin EM. Конститутивная экспрессия гена, ассоциированного с суточными часами 1 (CCA1), нарушает циркадные ритмы и подавляет собственную экспрессию. Клетка. 1998;93:1207–1217. [PubMed] [Google Scholar]
28. Якир Э., Хасидим М., Меламед-Бук Н., Хилман Д., Крон И., Грин Р.М. Автономные клеточные и специфичные для клеточного типа циркадные ритмы у арабидопсиса. Плант Дж. 2011; 68: 520–531. [PubMed] [Google Scholar]
29. Graf A., Coman D., Uhrig R.G., Walsh S., Flis A., Stitt M., Gruissem W. Параллельный анализ мутантов циркадных часов арабидопсиса выявил различные масштабы транскриптома и протеомная регуляция.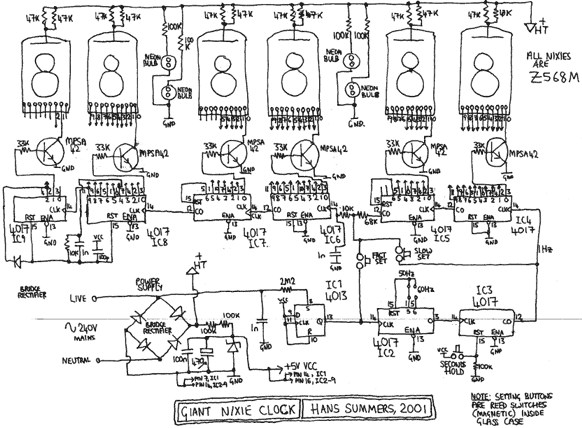 Открытая биол. 2017;7 [Бесплатная статья PMC] [PubMed] [Google Scholar]
Открытая биол. 2017;7 [Бесплатная статья PMC] [PubMed] [Google Scholar]
30. Крамер Дж., Хиндл М.М., Мартин С.Ф., Ле Бихан Т., Миллар А.Дж. Подготовка проб для фосфопротеомного анализа циркадных временных рядов у Arabidopsis thaliana. Методы Энзимол. 2015; 551:405–431. [Бесплатная статья PMC] [PubMed] [Google Scholar]
31. Элиас Дж. Э., Гиги С. П. Стратегия поиска мишени-приманки для протеомики на основе масс-спектрометрии. Методы Мол. биол. 2010; 604:55–71. [Бесплатная статья PMC] [PubMed] [Google Scholar]
qpMerge: слияние различных изоформ пептидов с использованием стратегии, ориентированной на мотив. bioRxiv. 2016 г.: 10.1101/047100. [препринт] [CrossRef] [Google Scholar]
33. Беренфаллер К., Хирш-Хоффманн М., Свозил Дж., Халл Р., Руссенбергер Д., Бишоф С., Лу К., Груиссем В., Багинский С. pep2pro: новый инструмент для комплексных данных о протеоме анализ для выявления информации об органоспецифических протеомах у Arabidopsis thaliana. интегр. биол. (Кэмб).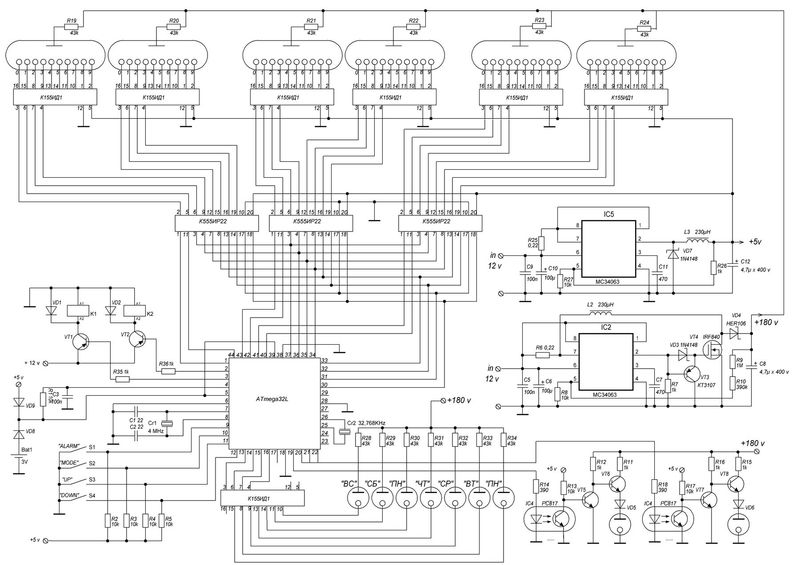 2011;3:225–237. [PubMed] [Google Scholar]
2011;3:225–237. [PubMed] [Google Scholar]
34. Vizcaíno J., Deutsch E., Wang R. ProteomeXchange обеспечивает глобально скоординированную отправку и распространение протеомных данных. Нац. Биотехнолог. 2014; 32: 223–226. [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Suzuki R., Shimodaira H. 2015. pvclust: Иерархическая кластеризация с P-значениями посредством многомасштабной бутстрепной передискретизации. [Google Scholar]
36. Хьюз М.Е., Хогенеш Дж.Б., Корнакер К. JTK_CYCLE: эффективный непараметрический алгоритм для обнаружения ритмических компонентов в наборах данных масштаба генома. Дж. Биол. Ритмы. 2010;25:372–380. [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Benjamini Y., Hochberg Y. Контроль частоты ложных открытий: практичный и мощный подход к множественному тестированию. Дж. Р. Стат. соц. Б. 1995;57:289–300. [Google Scholar]
38. Xue Y., Ren J., Gao X., Jin C., Wen L., Yao X. GPS 2.0, инструмент для прогнозирования специфических для киназы сайтов фосфорилирования в иерархии.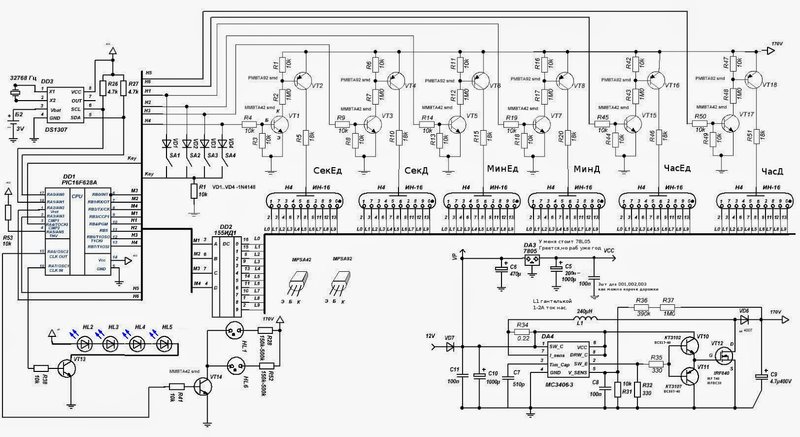 Мол. Клеточная протеомика. 2008; 7: 1598–1608. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Мол. Клеточная протеомика. 2008; 7: 1598–1608. [Бесплатная статья PMC] [PubMed] [Google Scholar]
39. Alexa A., Rahnenführer J., Lengauer T. Улучшенная оценка функциональных групп по данным экспрессии генов путем декорреляции структуры графа GO. Биоинформатика. 2006; 22:1600–1607. [PubMed] [Google Scholar]
40. Kalt-Torres W., Kerr P.S., Usuda H., Huber S.C. Суточные изменения в фотосинтезе листьев кукурузы: I. Скорость углеродного обмена, скорость экспорта ассимилятов и активность ферментов. Завод Физиол. 1987;83:283–288. [Статья бесплатно PMC] [PubMed] [Google Scholar]
41. Ван Шафтинген Э., Ледерер Б., Бартронс Р., Херс Х. Г. Кинетическое исследование пирофосфата: фруктозо-6-фосфатфосфотрансферазы из клубней картофеля. Приложение к микроанализу фруктозо-2,6-бисфосфата. Евро. Дж. Биохим. 1982; 129: 191–195. [PubMed] [Google Scholar]
42. Вилладсен Д., Нильсен Т.Х. Усечение N-конца влияет на кинетику и структуру фруктозо-6-фосфат-2-киназы/фруктозо-2,6-бисфосфатазы из Arabidopsis thaliana. Биохим. Дж. 2001; 359: 591–597. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Биохим. Дж. 2001; 359: 591–597. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Циркадная регуляция белка в зеленой линии I. Фосфорный рассвет предвосхищает начало света до пика белков в дневное время. bioRxiv. 2018 г.: 10.1101/287862. [препринт] [CrossRef] [Google Scholar]
44. Храбак Э.М., Чан К.В., Грибсков М., Харпер Дж.Ф., Чой Дж.Х., Халфорд Н., Кудла Дж., Луан С., Ниммо Х.Г., Суссман М.Р., Томас М. ., Уокер-Симмонс К., Чжу Дж. К., Хармон А. С. Надсемейство протеинкиназ CDPK-SnRK арабидопсиса. Завод Физиол. 2003; 132: 666–680. [Бесплатная статья PMC] [PubMed] [Google Scholar]
45. Wang Y., Liu Z., Cheng H., Gao T., Pan Z., Yang Q., Guo A., Xue Y. EKPD: Иерархическая база данных эукариотических протеинкиназ и протеинфосфатаз. Нуклеиновые Кислоты Res. 2014;42:D496–D502. [Бесплатная статья PMC] [PubMed] [Google Scholar]
46. Cho H.Y., Wen T.N., Wang Y.T., Shih M.C. Количественная фосфопротеомика протеинкиназы SnRK1, регулирующей фосфорилирование белков арабидопсиса при погружении в воду. Дж. Эксп. Бот. 2016;67:2745–2760. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Дж. Эксп. Бот. 2016;67:2745–2760. [Бесплатная статья PMC] [PubMed] [Google Scholar]
47. Нукаринен Э., Нэгеле Т., Педротти Л., Вурцингер Б., Майр А., Ландграф Р., Бёрнке Ф., Хэнсон Дж., Тейге М., Баэна-Гонсалес Э., Дроге-Лазер В. ., Weckwerth W. Количественная фосфопротеомика раскрывает роль растительного ортолога AMPK SnRK1 в качестве главного регулятора метаболизма при дефиците энергии. науч. Отчет 2016; 6: 31697. [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Сагден С., Донахи П.Г., Хэлфорд Н.Г., Харди Д.Г. Две родственные SNF1 протеинкиназы из листьев шпината фосфорилируют и инактивируют 3-гидрокси-3-метилглутарил-коэнзим А редуктазу, нитратредуктазу и сахарозофосфатсинтазу 9.0258 in vitro . Завод Физиол. 1999; 120: 257–274. [Бесплатная статья PMC] [PubMed] [Google Scholar]
49. Джамшир К. М., Лакшми А. Экспрессия FCS-подобных генов цинковых пальцев арабидопсиса по-разному регулируется сахарами, уровнем клеточной энергии и абиотическим стрессом.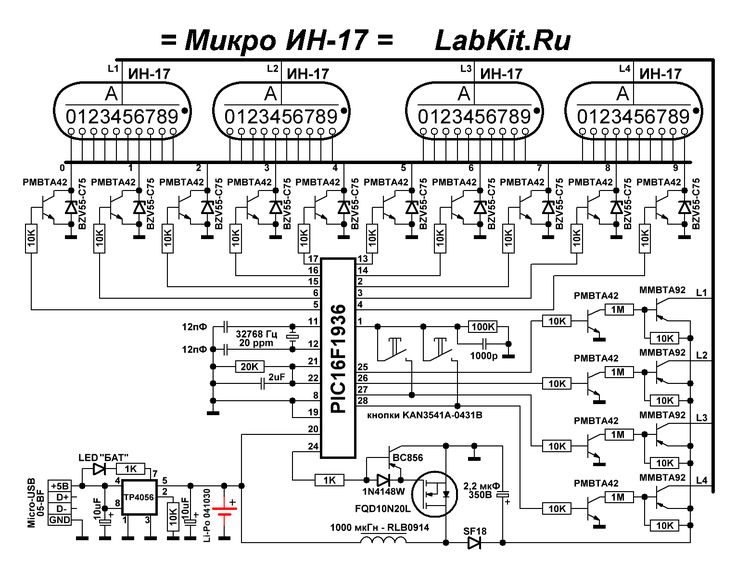 Фронт. Растениевод. 2015;6:746. [Статья бесплатно PMC] [PubMed] [Google Scholar]
Фронт. Растениевод. 2015;6:746. [Статья бесплатно PMC] [PubMed] [Google Scholar]
50. Jamsheer K M., Shukla B.N., Jindal S., Gopan N., Mannully CT, Laxmi A. FCS-подобный каркас цинковых пальцев киназы SnRK1 образованы скоординированными действиями домена FLZ и внутренне неупорядоченных областей. Дж. Биол. хим. 2018;293:13134–13150. [Бесплатная статья PMC] [PubMed] [Google Scholar]
51. Covington M.F., Maloof J.N., Straume M., Kay S.A., Harmer S.L. Анализ глобального транскриптома показывает циркадную регуляцию ключевых путей роста и развития растений. Геном биол. 2008;9:130 р. [Бесплатная статья PMC] [PubMed] [Google Scholar]
52. Liu X., Zhu Y., Zhai H., Cai H., Ji W., Luo X., Li J., Bai X. AtPP2CG1, a протеинфосфатаза 2С положительно регулирует солеустойчивость арабидопсиса зависимым от абсцизовой кислоты образом. Биохим. Биофиз. Рез. коммун. 2012; 422:710–715. [PubMed] [Академия Google]
53. Саху С.А., Рагхуванши Р., Шривастава А.К., Супрасанна П. В: Протеиновые фосфатазы и управление стрессом у растений. Пандей Г.К., редактор. Международное издательство Спрингер; 2020. Фосфатазы: критический регулятор устойчивости растений к абиотическим стрессам; стр. 163–201. [Google Scholar]
Пандей Г.К., редактор. Международное издательство Спрингер; 2020. Фосфатазы: критический регулятор устойчивости растений к абиотическим стрессам; стр. 163–201. [Google Scholar]
54. Ю Л.П., Миллер А.К., Кларк С.Е. POLTERGEIST кодирует протеинфосфатазу 2C, которая регулирует пути CLAVATA, контролирующие идентичность стволовых клеток в меристемах побегов и цветков арабидопсиса. Курс. биол. 2003;13:179–188. [PubMed] [Google Scholar]
55. Holtorf S., Apel K., Bohlmann H. Сравнение различных конститутивных и индуцибельных промоторов для сверхэкспрессии трансгенов в Arabidopsis thaliana. Завод Мол. биол. 1995; 29: 637–646. [PubMed] [Google Scholar]
56. Донахью Дж.Л., Алфорд С.Р., Торабинежад Дж., Кервин Р.Е., Нурбахш А., Рэй В.К., Херник М., Хуанг Х., Лайонс Б.М., Хейн П.П., Гилласпи Г.Э. Ген мио-инозитол-1-фосфатсинтазы1 Arabidopsis thaliana необходим для синтеза мио-инозитола и подавления гибели клеток. Растительная клетка. 2010; 22:888–903. [Бесплатная статья PMC] [PubMed] [Google Scholar]
57. Ma L., Tian T., Lin R., Deng X.W., Wang H., Li G. Arabidopsis FHY3 и FAR1 регулируют световые биосинтез инозитола и ответы на окислительный стресс путем транскрипционной активации MIPS1. Мол. Завод. 2016; 9: 541–557. [PubMed] [Google Scholar]
Ma L., Tian T., Lin R., Deng X.W., Wang H., Li G. Arabidopsis FHY3 и FAR1 регулируют световые биосинтез инозитола и ответы на окислительный стресс путем транскрипционной активации MIPS1. Мол. Завод. 2016; 9: 541–557. [PubMed] [Google Scholar]
58. Эдвардс К.Д., Андерсон П.Е., Холл А., Салатия Н.С., Локк Дж.К., Линн Дж.Р., Страуме М., Смит Дж.К., Миллар А.Дж. FLOWERING LOCUS C опосредует естественные колебания в высокотемпературной реакции циркадных часов арабидопсиса. Растительная клетка. 2006;18:639–650. [Бесплатная статья PMC] [PubMed] [Google Scholar]
59. Covington M.F., Harmer S.L. Циркадные часы регулируют передачу сигналов ауксина и ответы арабидопсиса. PLoS биол. 2007; 5: 1773–1784. [Бесплатная статья PMC] [PubMed] [Google Scholar]
60. Монро Дж. Д. Участие пяти каталитически активных β-амилаз арабидопсиса в метаболизме крахмала листьев и росте растений. Завод Директ. 2020;4 [Бесплатная статья PMC] [PubMed] [Google Scholar]
61. Граф А., Шлерет А. , Ститт М., Смит А.М. Циркадный контроль доступности углеводов для роста растений арабидопсиса в ночное время. проц. Натл. акад. науч. США 2010; 107:9458–9463. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Ститт М., Смит А.М. Циркадный контроль доступности углеводов для роста растений арабидопсиса в ночное время. проц. Натл. акад. науч. США 2010; 107:9458–9463. [Бесплатная статья PMC] [PubMed] [Google Scholar]
62. Smith S.M., Fulton D.C., Chia T., Thorneycroft D., Chapple A., Dunstan H., Hylton C., Zeeman S.C., Smith A.M. Суточные изменения транскриптома, кодирующего ферменты метаболизма крахмала, свидетельствуют как о транскрипционной, так и о посттранскрипционной регуляции метаболизма крахмала в листьях арабидопсиса. Завод Физиол. 2004; 136: 2687–2699. [Бесплатная статья PMC] [PubMed] [Google Scholar]
63. Bläsing O.E., Gibon Y., Günther M., Höhne M., Morcuende R., Osuna D., Thimm O., Usadel B., Scheible W.R., Ститт М. Сахара и циркадная регуляция вносят большой вклад в глобальную регуляцию суточной экспрессии генов у арабидопсиса. Растительная клетка. 2005;17:3257–3281. [Бесплатная статья PMC] [PubMed] [Google Scholar]
64. Коруцци Г.М. Первичная ассимиляция N в аминокислоты у арабидопсиса. араб. B. 2003;2 [бесплатная статья PMC] [PubMed] [Google Scholar]
араб. B. 2003;2 [бесплатная статья PMC] [PubMed] [Google Scholar]
65. Wong H.K., Chan H.K., Coruzzi G.M., Lam H.M. Корреляция экспрессии гена ASN2 с метаболизмом аммония у арабидопсиса. Завод Физиол. 2004; 134:332–338. [PMC free article] [PubMed] [Google Scholar]
66. Карран А., Чанг И.Ф., Чанг К.Л., Гарг С., Мигель Р.М., Бэррон Ю.Д., Ли Ю., Романовский С., Кушман Дж.К., Грибсков М. , Harmon A.C., Harper J.F. Кальций-зависимые протеинкиназы из арабидопсиса демонстрируют различия в субстратной специфичности при анализе 103 субстратов. Фронт. Растениевод. 2011;2:36. [Бесплатная статья PMC] [PubMed] [Google Scholar]
67. Клайн К.Г., Барретт-Уилт Г.А., Суссман М.Р. Изменения фосфорилирования белка в растениях, вызванные растительным гормоном абсцизовой кислотой. проц. Натл. акад. науч. США 2010;107:15986–15991. [Бесплатная статья PMC] [PubMed] [Google Scholar]
68. Pou A., Jeanguenin L., Milhiet T., Batoko H., Chaumont F., Hachez C. Опосредованная соленостью транскрипционная и посттрансляционная регуляция Аквапорин PIP2;7 арабидопсиса. Завод Мол. биол. 2016;92:731–744. [PubMed] [Академия Google]
Завод Мол. биол. 2016;92:731–744. [PubMed] [Академия Google]
69. Prado K., Cotelle V., Li G., Bellati J., Tang N., Tournaire-Roux C., Martinière A., Santoni V., Maurel C. Осциллирующее фосфорилирование аквапоринов и белки 14-3-3 опосредуют циркадную регуляцию гидравлики листа. Растительная клетка. 2019; 31: 417–429. [Бесплатная статья PMC] [PubMed] [Google Scholar]
70. Chen L.Q., Qu X.Q., Hou B.H., Sosso D., Osorio S., Fernie A.R., Frommer W.B. Отток сахарозы, опосредованный белками SWEET, как ключевой этап транспорта флоэмы. Наука. 2012;335:207–211. [PubMed] [Академия Google]
71. Дейхолос М.К., Каванесс Г.Ф., Холл Б., Кинг Э., Пунвани Дж., Ван Норман Дж., Зибурт Л.Е. ВАРИКОЗ, белок с доменом WD, необходим для развития листовой пластинки. Разработка. 2003; 130:6577–6588. [PubMed] [Google Scholar]
72. Кава Д., Мейер А.Дж., Деккер Х.Л., Абд-Эль-Халием А.М., Геварт К., Ван Де Слейке Э., Машковска Дж., Бухолк М., Добровольская Г., Де Джегер Г., Шууринк Р. К., Харинг М.А., Тестеринк С. Протеинкиназы SnRK2 и механизм удаления мРНК контролируют развитие корней и реакцию на соль. Завод Физиол. 2020; 182: 361–377. [Бесплатная статья PMC] [PubMed] [Google Scholar]
К., Харинг М.А., Тестеринк С. Протеинкиназы SnRK2 и механизм удаления мРНК контролируют развитие корней и реакцию на соль. Завод Физиол. 2020; 182: 361–377. [Бесплатная статья PMC] [PubMed] [Google Scholar]
73. Нильсен Т.Х., Рунг Дж.Х., Вилладсен Д. Фруктоза-2,6-бисфосфат: светофор в метаболизме растений. Тенденции Растениевод. 2004; 9: 556–563. [PubMed] [Google Scholar]
74. Маккормик А.Дж., Крюгер Н.Дж. Недостаток фруктозо-2,6-бисфосфата ставит под угрозу фотосинтез и рост арабидопсиса в меняющихся условиях. Плант Дж. 2015; 81: 670–683. [PubMed] [Google Scholar]
75. Вилладсен Д., Рунг Дж.Х., Драборг Х., Нильсен Т.Х. Структура и гетерологичная экспрессия гена, кодирующего фруктозо-6-фосфат, 2-киназу/фруктозо-2,6-бисфосфатазу из Arabidopsis thaliana. Биохим. Биофиз. Акта. 2000;1492: 406–413. [PubMed] [Google Scholar]
76. Кулма А., Вилладсен Д., Кэмпбелл Д.Г., Мик С.Е., Хартхилл Дж.Е., Нильсен Т.Х., Маккинтош С. Фосфорилирование и 14-3-3 связывание 6-фосфофрукто-2- арабидопсиса киназа/фруктозо-2,6-бисфосфатаза.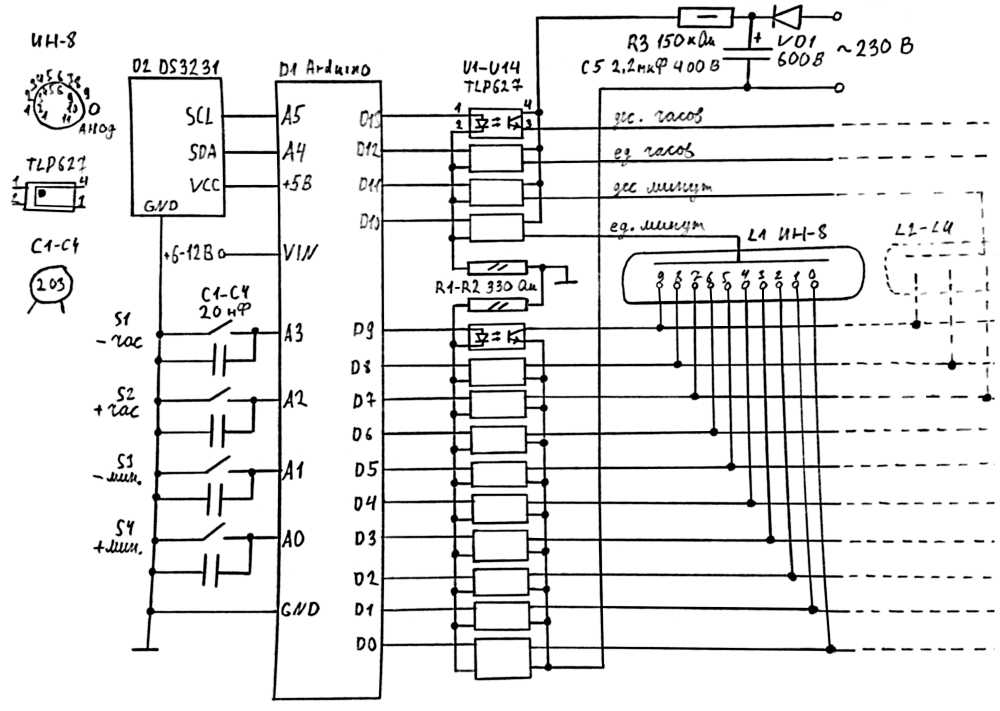 Плант Дж. 2004; 37: 654–667. [PubMed] [Google Scholar]
Плант Дж. 2004; 37: 654–667. [PubMed] [Google Scholar]
77. Bläsing O.E., Gibon Y., Günther M., Höhne M., Morcuende R., Osuna D., Thimm O., Usadel B., Scheible W.R., Stitt M. Sugars and циркадная регуляция вносит большой вклад в глобальную регуляцию суточной экспрессии генов у арабидопсиса. Растительная клетка. 2005;17:3257–3281. [Бесплатная статья PMC] [PubMed] [Google Scholar]
78. Sugano S., Andronis C., Ong M.S., Green R.M., Tobin E.M. Протеинкиназа CK2 участвует в регуляции циркадианных ритмов у арабидопсиса. проц. Натл. акад. науч. США, 1999; 96:12362–12366. [Бесплатная статья PMC] [PubMed] [Google Scholar]
79. Акаши М., Нисида Э. Участие киназного каскада MAP в сбросе циркадных часов млекопитающих. Гены Дев. 2000; 14: 645–649. [Бесплатная статья PMC] [PubMed] [Google Scholar]
80. Иитака С., Миядзаки К., Акаике Т., Исида Н. Роль гликогенсинтазкиназы-3бета в циркадных часах млекопитающих. Дж. Биол. хим. 2005;280:29397–29402. [PubMed] [Google Scholar]
81. Besing R.C., Paul J.R., Hablitz L.M., Rogers C.O., Johnson R.L., Young M.E., Gamble K.L. Суточная ритмичность активных изоформ GSK3 модулирует ритмы генов молекулярных часов в супрахиазматическом ядре. Дж. Биол. Ритмы. 2015;30:155–160. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Besing R.C., Paul J.R., Hablitz L.M., Rogers C.O., Johnson R.L., Young M.E., Gamble K.L. Суточная ритмичность активных изоформ GSK3 модулирует ритмы генов молекулярных часов в супрахиазматическом ядре. Дж. Биол. Ритмы. 2015;30:155–160. [Бесплатная статья PMC] [PubMed] [Google Scholar]
82. Мартинек С., Иноног С., Манукян А. С., Янг М. В. Роль гена полярности сегмента shaggy/GSK-3 в циркадных часах дрозофилы. Клетка. 2001;105:769–779. [PubMed] [Google Scholar]
83. Nietzsche M., Schießl I., Börnke F. Комплекс усложняется: белок-белковые взаимодействия SnRK1 с белками семейства DUF581 обеспечивают каркас для SnRK1, специфичных для клеток и типов стимулов. сигнализация у растений. Фронт. Растениевод. 2014;5:54. [Бесплатная статья PMC] [PubMed] [Google Scholar]
84. Shin J., Sánchez-Villarreal A., Davis AM, Du S.-x., Berendzen K.W., Koncz C., Ding Z., Li C. , Дэвис С.Дж. Метаболический датчик AKIN10 модулирует циркадные часы Arabidopsis в зависимости от света. Окружающая среда растительной клетки. 2017;40:997–1008. [PubMed] [Google Scholar]
2017;40:997–1008. [PubMed] [Google Scholar]
85. Санчес-Вильярреал А., Дэвис А.М., Дэвис С.Дж. Активность AKIN10 как клеточная связь между метаболизмом и включением циркадных часов у Arabidopsis thaliana. Сигнал завода. Поведение 2018;13 [бесплатная статья PMC] [PubMed] [Google Scholar]
86. Франк А., Матиолли К.С., Виана А.Дж.С., Хирн Т.Дж., Кусакина Дж., Белбин Ф.Е., Уэллс Ньюман Д., Йотикава А., Кано- Рамирез Д.Л., Чембат А., Крэгг-Барбер К., Хейдон М.Дж., Хотта С.Т., Винсенц М., Уэбб А.А.Р. и др. Циркадный захват у арабидопсиса чувствительным к сахару транскрипционным фактором bZIP63. Курс. биол. 2018;28:2597–2606.e6. [Бесплатная статья PMC] [PubMed] [Google Scholar]
87. Гонсалес А., Холл М.Н., Лин С.С., Харди Д.Г. AMPK и TOR: Инь и Ян клеточного восприятия питательных веществ и контроля роста. Клеточный метаб. 2020; 31: 472–492. [PubMed] [Google Scholar]
88. Ларкин М.А., Блэкшилдс Г., Браун Н.П., Ченна Р., Макгеттиган П.А., Маквильям Х., Валентин Ф.