
Схема электронных часов-будильника » Паятель.Ру
Категория: Часы
Электронные часы построены на основе комплекта микросхем К176ИЕ12, К176ИЕ13 и К176ИД2. Микросхемы очень хорошо работают с светодиодными семисегментными индикаторами, которые, хотя и недешевы, но отличаются высокой надежностью, и что самое главное не раздражают зрение как электролюминесцетные. Часы-будильник отсчитывают текущее время в часах и минутах, секунды индицируются одним светодиодом, который на табло размещается между разрядами часов и минут. Время будильника устанавливается в течении суток. При совпадении времени будильника и текущего времени включается музыкальный сигнал, один из восьми, который можно выбрать заранее.
Кроме срабатывания звукового сигнала часы-будильник могут включать любую нагрузку, питающуюся напряжением до 220В и имеющую мощность не более 100 Вт, это может быть радиоприемник, магнитофон, любой электроприбор не превышающий эту мощность.
Часы питаются от сети переменного тока 220В и от резервного источника на 9В (батарея типа «Кроны»). При отключении электроснабжения часы переходят на питание от резервного источника, при этом индикация, с целью экономии электроэнергии, отключается, а все остальные функции, включая и будильник (кроме выключателя электроприбора) сохраняются. Таким образом, даже если электричество отключили на сутки будильник прозвучит вовремя.
Еще одно преимущество — громкость будильника настолько высока, что разбудит даже самого крепко спящего человека.
Принципиальная схема показана на рисунке. Собственно часы-будильник собраны на микросхемах D1-D3 по упрощенной типовой схеме. Микросхема D1 К176ИЕ12 представляет собой формирователь временных последовательностей. Она содержит мультивибратор с кварцевым резонатором Q1 и два счетчика. Первый вырабатывает секундные и полсекундные импульсы, а также сдвинутые по фазе на четверть периода импульсы частотой 128 Гц для реализации динамической индикации. Второй счетчик имеет коэффициент пересчета 60 и служит для формирования минутных импульсов.
Второй счетчик имеет коэффициент пересчета 60 и служит для формирования минутных импульсов.
Микросхема D2 содержит счетчики часов и минут, а также ОЗУ будильника и формирователь сигнала будильника. Как только информация записанная в ОЗУ совпадает с текущем времени на выводе 7 D2 появляются пачки импульсов, которые в типовом включении должны поступать на пьезоизлучатель.
Установка текущего времени и будильника выполняется тремя кнопками S1-S3. При нажатии на S1 показания минут увеличиваются с частотой 2 Гц, при нажатии на S2 тоже самое происходит с показаниями часов. При нажатии S3 на индикацию выводится время установки будильника и при этом можно кнопками S1 и S2 установить время будильника.
D3 — дешифратор для индикаторов с семи-сегментным формированием цифр. Двоичный четырехразрядный код каждой цифры поступает на его входы «1-2-4-8», при этом смена данных в соответствии с динамической индикацией производится импульсом, поступающим на его вывод 1.
На выходе дешифратора включены четыре семисегментных индикатора АЛС324Б. Одинаковые катодные выводы сегментов этих индикаторов соединены вместе. Таким образом, что один и тот же код поступает сразу на катоды всех четырех индикаторов. Выбор какой из индикаторов при этом будет светиться производится при помощи четырех транзисторных ключей VT2-VT5, на базы которых поступают импульсные последовательности частотой 128 Гц и скважностью 4, сдвинутые по фазе относительно друг друга на четверть периода.
Одинаковые катодные выводы сегментов этих индикаторов соединены вместе. Таким образом, что один и тот же код поступает сразу на катоды всех четырех индикаторов. Выбор какой из индикаторов при этом будет светиться производится при помощи четырех транзисторных ключей VT2-VT5, на базы которых поступают импульсные последовательности частотой 128 Гц и скважностью 4, сдвинутые по фазе относительно друг друга на четверть периода.
Сигнал будильника выпрямляется диодом VD8 и на конденсаторе С10 появляется некоторое напряжение, которое поступает на транзисторный ключ на VT8 и открывает его. При этом ток протекает через обмотку электромагнитного реле Р1, его контакты приходят в движение и замыкают цепь нагрузки «Н» и цепь «коллектор-эмиттер» транзистора VT8. Таким образом реле самоблокируется и остается во включенном состоянии даже тогда, когда сигнал будильника прекращается. Вывести реле из этого положения можно только кратковременным прекращением тока через него при помощи размыкающей кнопки S6.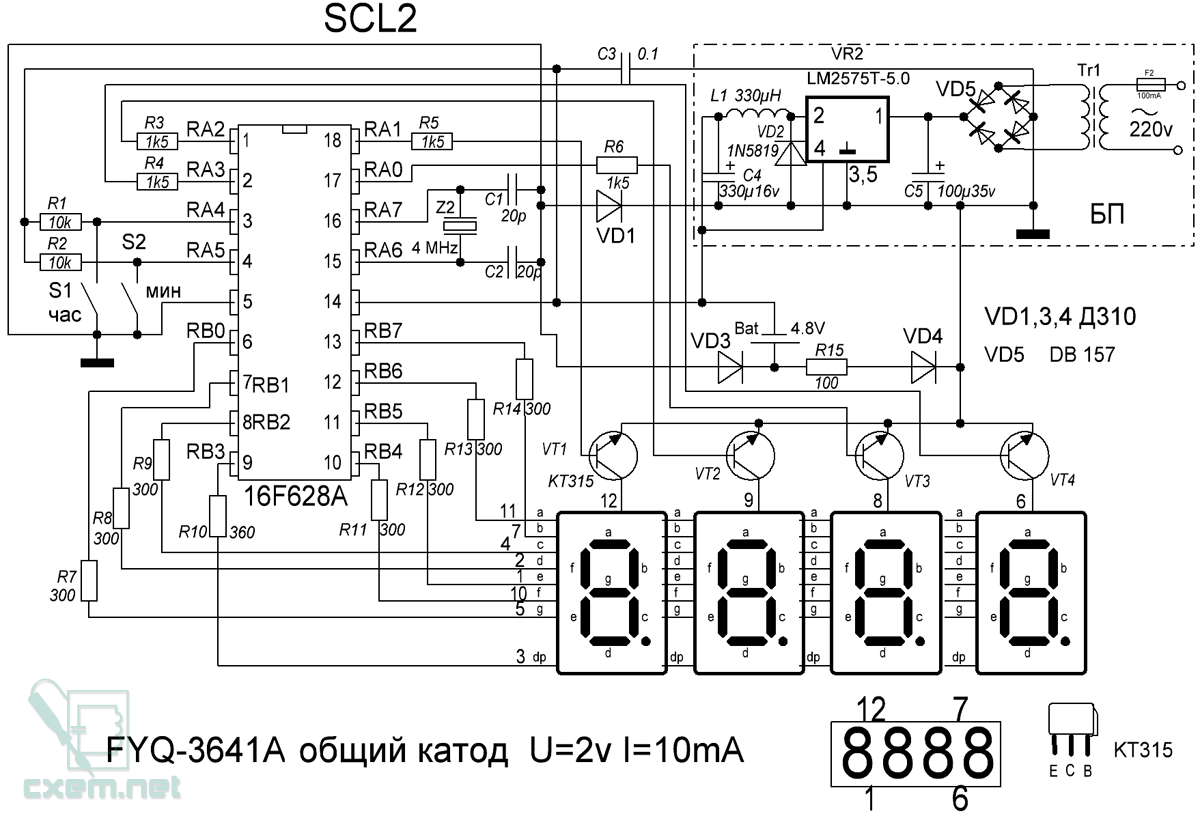
Одновременно импульсы будильника поступают через конденсатор С7 на вход запуска (вывод 13) универсального музыкального синтезатора УМС8-08. Синтезатор запускается и импульсный сигнал с его выхода (вывод 1) поступает на транзисторный импульсный усилитель на VT6 и VT7. В коллекторной цепи VT7 включен динамический громкоговоритель.
В результате такого построения выходного каскада громкость звучания получается достаточно большой. При необходимости её снизить можно включением резистора на 20-100 Ом последовательно с динамиком. Отключается будильник при помощи выключателя SB1, который отключает динамик.
Питается УМС от параметрического стабилизатора на R18 и VD5. Тактовая частота на него поступает с вывода 14 D1 через С5. Кнопка S4 служит для ручного запуска синтезатора, a S5 для выбора мелодии которая должна будет исполняться.
Сетевой источник питания сделан на маломощном силовом трансформаторе Т1. Постоянное напряжение 12-15В с выхода мостового выпрямителя питает электромагнитное реле Р1. Схема часов питается от стабилизатора на VT9, вырабатывающего 10В. Резервный источник G1 подключается через диод VD6. Когда сетевое напряжение есть он защищен этим диодом и не используется. При отключении сетевого напряжения диод VD7 препятствует поступлению тока от G1 на светодиодные индикаторы.
Схема часов питается от стабилизатора на VT9, вырабатывающего 10В. Резервный источник G1 подключается через диод VD6. Когда сетевое напряжение есть он защищен этим диодом и не используется. При отключении сетевого напряжения диод VD7 препятствует поступлению тока от G1 на светодиодные индикаторы.
Светодиодные индикаторы АЛС324Б можно заменить на любые другие семисегментные с общим анодом. Электромагнитное реле Р1 типа РЭС22 на напряжение 10-15В. Синтезатор УМС8-08 можно заменить на любой другой УМС из серии УМС8 и УМС7. Все транзисторы могут быть с любыми буквенными индексами. КТ 315 желательно заменить на КТ503.
Трансформатор Т1 готовый, на его вторичной обмотке имеется переменное напряжение 12В. Можно взять любой другой трансформатор с выходным напряжением 12-22В, при этом соответственно придется выбрать С11 на другое напряжение и реле, либо подключить реле через гасящий резистор.
Динамик любого типа, например от малогабаритного радиоприемника.
часы — радиоэлектроника, схемы и статьи
Ниже приведены принципиальные схемы и статьи по тематике «часы» на сайте по радиоэлектронике и радиохобби RadioStorage. net .
net .
Что такое «часы» и где это применяется, принципиальные схемы самодельных устройств которые касаются термина «часы».
1)Часы для автоматического управления устройствами (537РУ2, 176ИЕ12)
Для автоматического управления режимом работы различных бытовых электроприборов или радиоаппаратуры в домашних условиях, а также на производстве иногда бывает необходимо иметь время-задающий автомат. Например, такое устройство может по заданной программе управлять поливом растений …
2)Таймер из электронных часов ЦАТ-01
Электронные цифровые часы-будильник на базе микроконтроллера КА1035ХЛ1 выпускались в середине 1990 г. Ленинградским объединением «Светлана» и в свое время наводнили рынок. Прибор установливается в автомобиле для индикации времени и имеет функцию программируемого…
3)Солнечные часы
Время быстротечно, и мы не можем управлять им. Тысячелетия человек пытается остановить время, но, увы, в результате лишь наблюдает его ход. Солнечные часы — древнейший инструмент, позволяющий следить за ходом времени. Их использовали…
Солнечные часы — древнейший инструмент, позволяющий следить за ходом времени. Их использовали…
4)Коммутатор для часов на микросхеме К145ИК1901
Лет 10-15 назад промышленностью выпускался электронный конструктор под названием “Старт 7178”, предназначенный для самостоятельной сборки часов. В этот набор входил полный комплект радиодеталей для…
5)Электронные часы с бестрансформаторным питанием от сети
Особенности этих часов — бестрансформаторное питание и возможность применения в них кварцевого резонатора на любую частоту от 50 до 560 кГц. На микросхеме А1 собран кварцевый генератор с резонатором Z1. Элементы D1.1 и D2.1 формируют из синусоидального напряжения прямоугольные импульсы,…
6)Питание часов-будильника 1,5В от автомобильной бортовой сети
Карманные часы-будильник Miracle или аналогичные, питаются обычно от одного гальванического элемента напряжением 1,5V. При желании установить их в автомобиле есть только один приемлемый способ крепления — приклеить их двухсторонним скотчем к торпеде автомобиля в удобном месте. Однако, в таком …
При желании установить их в автомобиле есть только один приемлемый способ крепления — приклеить их двухсторонним скотчем к торпеде автомобиля в удобном месте. Однако, в таком …
7)Огромный циферблат из светодиодных лент, часы на микросхемах (IRLU024N)
При оформлении одного мероприятия потребовалось сделать цифровые часы сгигантским дисплеем высотой цифр по два метра. Самое сложное в данном процессе было выбрать каким образом и из чего сделать этот самый дисплей. К счастью, сейчас продаются светодиодные ленты. Было решено сделать дисплей с …
8)Часы на люминесцентных индикаторах ИВ-11 (К176ИЕ18, К176ИЕ13)
9)Цифровые часы на микросхемах HCF4521, HCF4026BEY
Лет 25 тому назад простые цифровые часы на «россыпи» делали используя специализированные микросхемы серии К176.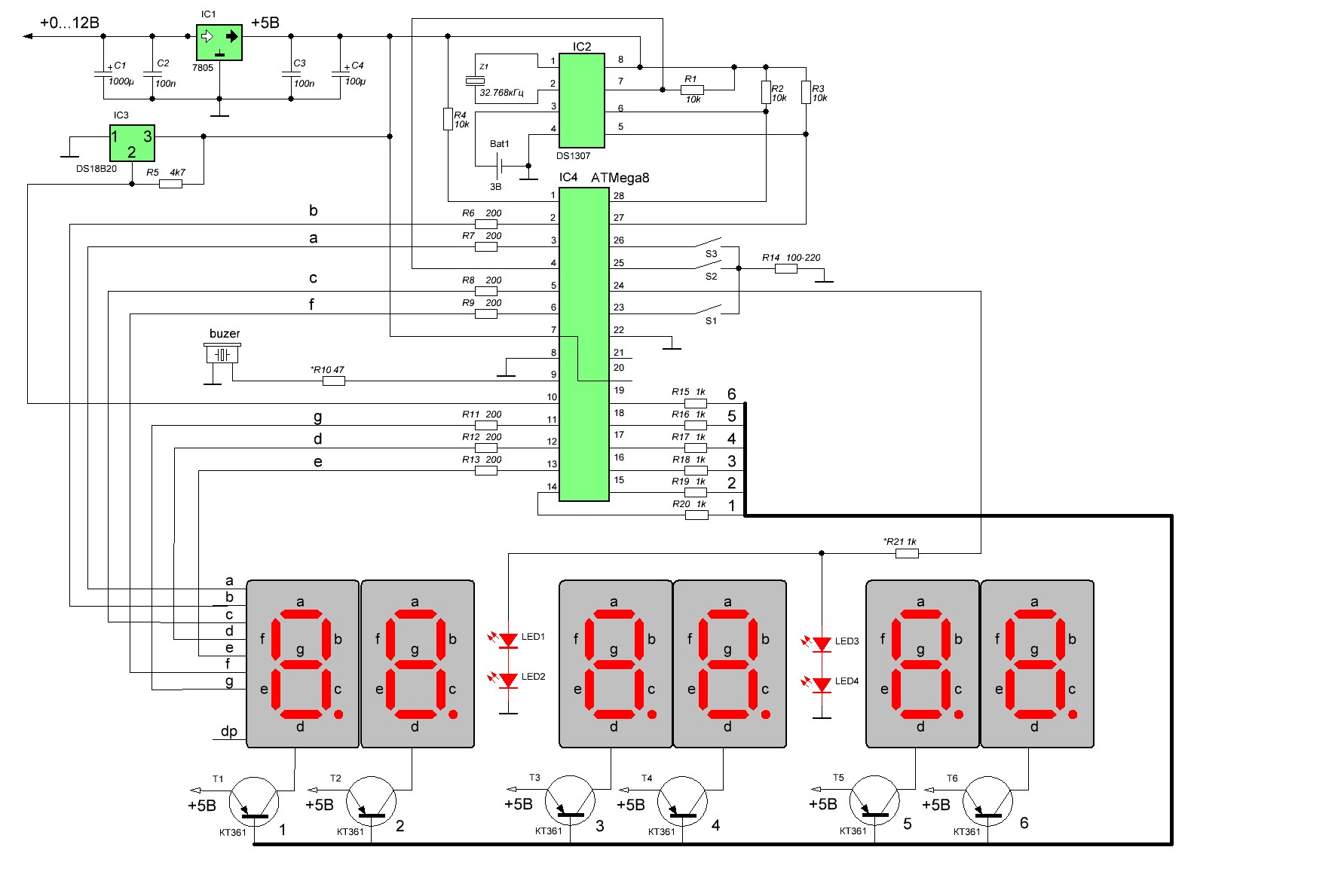 Даже продавались наборы -конструкторы для сборки часов со статической индикацией, обычно, на микросхемах К176ИЕ5, К176ИЕЗ, К176ИЕ4. Сейчас эти микросхемы уже давно не …
Даже продавались наборы -конструкторы для сборки часов со статической индикацией, обычно, на микросхемах К176ИЕ5, К176ИЕЗ, К176ИЕ4. Сейчас эти микросхемы уже давно не …
10)Часы с термометром и таймером на PIC-микроконтроллере PIC16F873A
Предлагаемое устройство отсчитывает время, измеряет температуру в доме и на улице, выключает по истечении заданного времени сетевую нагрузку. Оно просто в изготовлении. Вся информация выводится на светодиодный индикатор, который видно как днем, так и ночью, как вблизи, так и издалека. В продаже …
11)Простой стабилизатор напряжения для часов на +1,5В
Принципиальная схема очень простого стабилизатора напряжения на 1,5В для питания настенных часов от зарядного устройства USB 5V. В последнее время цена универсального зарядного устройства с USB разъемом на выходе сильно снизилась, и доходит до величины менее 1$. При том что это вполне …
12)Цифровые электронные часы на индикаторах ИН-12 или ИН-18 (К176ИЕ12, К561ИЕ8)
Схема и описание цифровых электронных часов на индикаторах ИН-12(ИН-18) и микросхемах К176ИЕ12, К561ИЕ8.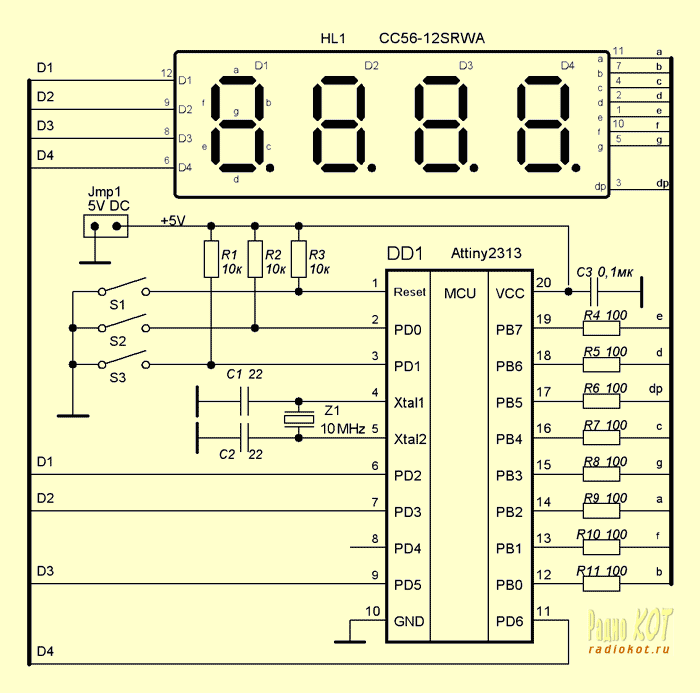 В настоящее на просторах интернета можно встретить множество всевозможных схем и конструкций часов на микроконтроллерах и практически уже нет схем на обычной логической элементной базе. Я нашел только три подробных схемы электронных часов на логических микросхемах. Схемы устройств на основе микроконтроллеров, можно сказать, по всем параметрам выигрывают у старых схем на обычной элементарной базе …
В настоящее на просторах интернета можно встретить множество всевозможных схем и конструкций часов на микроконтроллерах и практически уже нет схем на обычной логической элементной базе. Я нашел только три подробных схемы электронных часов на логических микросхемах. Схемы устройств на основе микроконтроллеров, можно сказать, по всем параметрам выигрывают у старых схем на обычной элементарной базе …
13)Часы-будильник на микроконтроллере АТ89С2051 и DS1307
Схема самодельных часов-будильника для установки в приборную панель автомобиля. Устройство питаются от автомобильного аккумулятора. Однако, они с таким же успехом могут использоваться и дома. В этом случае просто отсутствует подсветка дисплея, а питание осуществляется от любого сетевого …
14)Самодельные цифровые часы с индикаторами из светодиодных лент (К176ИЕ12, К176ИЕ4)
Для установки на проходных предприятий, вокзалах, в торговых центрах и в других местах массового прохода людей необходимы электронные часы с очень крупным и ярким дисплеем.
15)Сетевой блок питания на 1,5В для электромеханических часов
Электромеханические часы обычно питаются от элемента на 1,5V. Его можно заменить сетевым источником, схема которого показана здесь. В ней в качестве стабилитрона используется ИК-светодиод с прямым напряжением около 1,5V. Механизм часов питается от этого напряжения. Рис. 1. Схема сетевого …
16)Часы с календарем на индикаторах ИН-12
Предлагаемые часы показывают текущее время и дату, обладают функциями будильника. Их особенность — использование газоразрядных цифровых индикаторов ИН-12. Подобные индикаторы широко применялись в электронных часах и цифровых измерительных приборах в семидесятые годы прошлого века. Индикаторы …
17)Двоичные часы на микроконтроллере PIC16F628A
Схема самодельных двоичных часов на микроконтроллере PIC16F628A и светодиодах. Эти необычные карманные часы могут стать оригинальным подарком. Индикатор времени в них построен всего на шести единичных светодиодах. Секрет в том, что число часов и число минут текущего времени отображаются ими …
Эти необычные карманные часы могут стать оригинальным подарком. Индикатор времени в них построен всего на шести единичных светодиодах. Секрет в том, что число часов и число минут текущего времени отображаются ими …
18)Блок питания для кварцевых часов на ионисторе (LM317)
Сейчас продается очень много различных настенных и настольных кварцевых часов довольно больших габаритов. Обычно, там пустой корпус, а посредине его размещен типовой пластмассовый механизм, вроде того, что в кварцевых малогабаритных будильниках. Разница только в том, что стрелки больше …
Схема энергосберегающих тактовых импульсов в новых ПК
Давно изученная стратегия повторного использования энергии, используемой для тактирования компьютерных процессоров, теперь начинает внедряться в коммерческие чипы. Подход, получивший название резонансного тактирования, был интегрирован в новый процессор Advanced Micro Devices AMD A10-4600M. Чип, известный под кодовым названием Trinity, дебютировал в мае и сейчас поставляется в настольных и портативных компьютерах.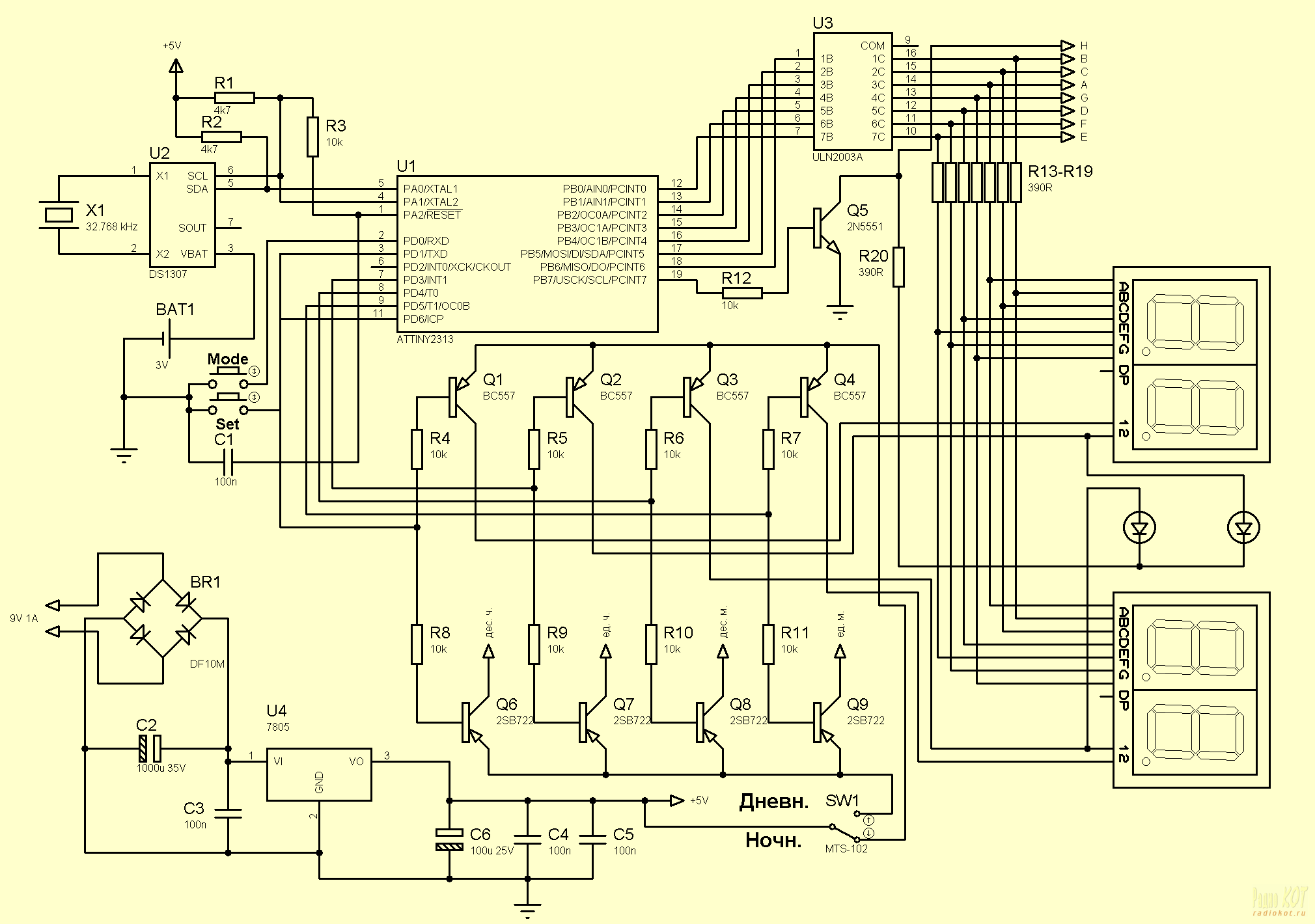
Следуя примеру маятника, технология использует катушки индуктивности для периодического накопления и высвобождения энергии, которая обычно рассеивается через сеть распределения часов на микросхеме. По данным AMD и Cyclos Semiconductor, которые предоставили процессорной фирме лицензию на эту технологию, такой подход может снизить мощность, необходимую для тактовой частоты чипов, на 24%. В качестве альтернативы, эту технологию можно использовать для повышения максимальной скорости чипов с поддержкой 4 гигагерц на 100 мегагерц, не требуя дополнительной мощности.
Найти способы экономии тактовой мощности, которая может потреблять 30 % энергии высокопроизводительного чипа, непросто. Чипам необходимо стабильное сердцебиение часов для управления вычислениями. И сигналы часов должны быть широко распространены, в десятках тысяч или даже в сотнях тысяч мест.
Производители микросхем традиционно использовали «деревья» для распределения тактовых сигналов. В них используется множество ответвляющихся проводов и сетей небольших цепей для приема и распространения тактового сигнала.
Разработчики высокопроизводительных чипов в значительной степени решили эту проблему, добавив проводную сетку, часто называемую сеткой часов, которая закорачивает конечные точки дерева часов и, таким образом, минимизирует колебания синхронизации. Но такой подход достигается за счет мощности. Сетки часов — это, по сути, большие конденсаторы; впрыскивание и протягивание заряда через весь добавленный металл потребляет много энергии.
Около 10 лет назад исследователи начали искать, казалось бы, простое решение: подключить к сетке несколько катушек индуктивности.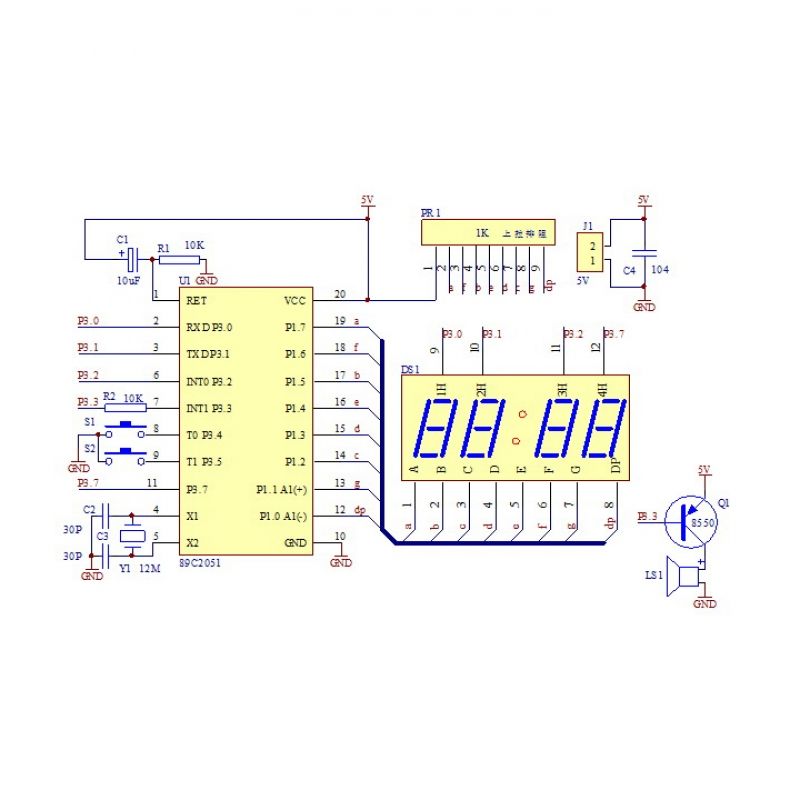 В результате получается LC-генератор, который естественным образом колеблется между двумя напряжениями, поскольку он попеременно накапливает энергию в сетке и в катушках индуктивности. Микросхема, использующая такой генератор для создания «резонансных часов», может полагаться на меньшие по размеру и менее энергоемкие схемы для управления своей сетью распределения часов. Они понадобятся только для запуска генератора и подачи небольшого количества энергии во время каждого цикла, чтобы поддерживать его работу, как толкание качелей.
В результате получается LC-генератор, который естественным образом колеблется между двумя напряжениями, поскольку он попеременно накапливает энергию в сетке и в катушках индуктивности. Микросхема, использующая такой генератор для создания «резонансных часов», может полагаться на меньшие по размеру и менее энергоемкие схемы для управления своей сетью распределения часов. Они понадобятся только для запуска генератора и подачи небольшого количества энергии во время каждого цикла, чтобы поддерживать его работу, как толкание качелей.
Cyclos, стартап, базирующийся в Беркли, Калифорния, и Анн-Арборе, Мичиган, находился в скрытом режиме с 2009 года, когда компания сообщила о проверке принципа действия, показывающем, что она может использовать один индуктор, расположенный вне чипа, для восстановления до 85 процентов энергии в сети распределения тактовых импульсов чипа микроконтроллера ARM с частотой 200 МГц.
Полная интеграция технологии в высокопроизводительный чип сопряжена с рядом проблем.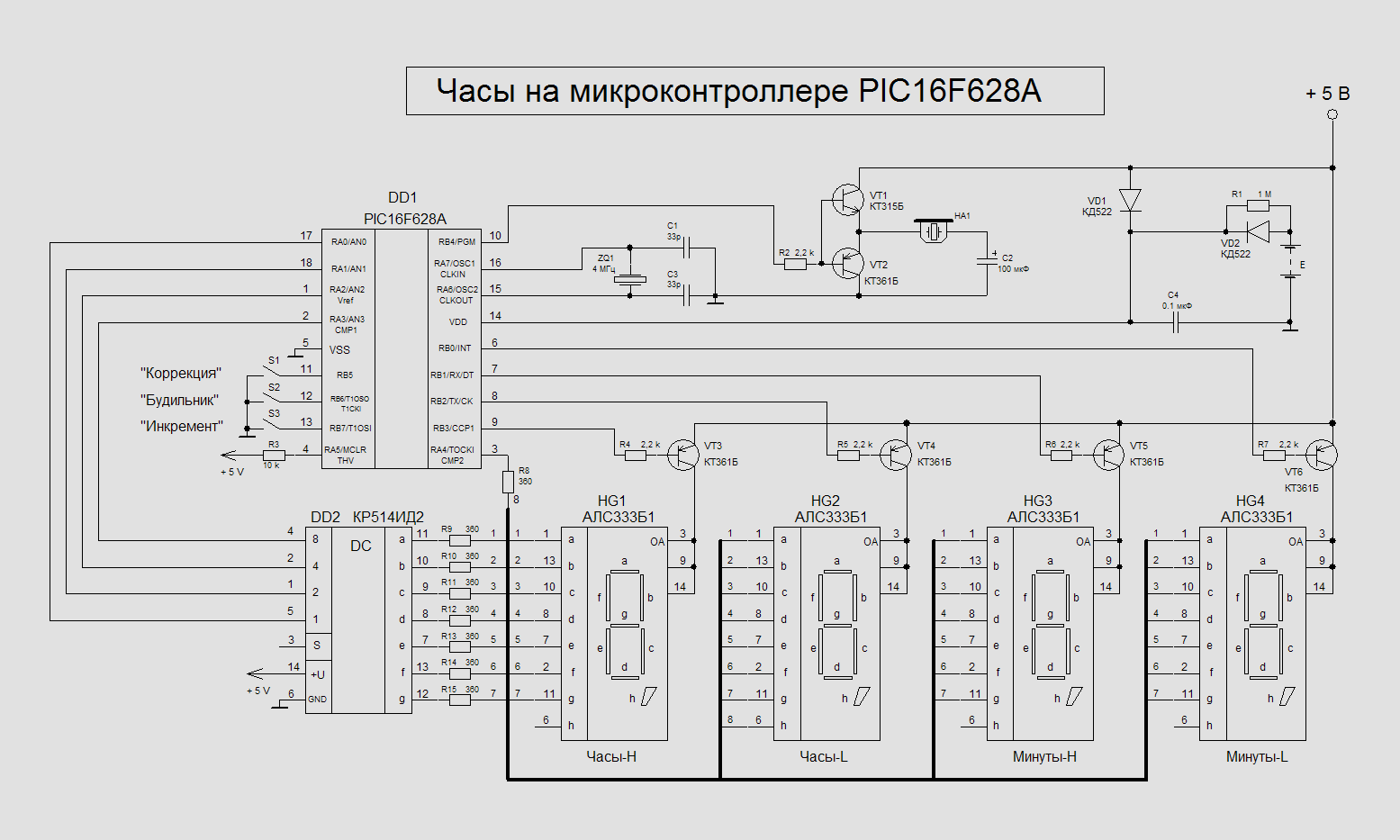 Во-первых, говорит соучредитель Cyclos Мариос С. Папаефтимиу, профессор электротехники и компьютерных наук в Мичиганском университете, команда должна была найти способ включить спиральные петли катушек индуктивности в и без того довольно плотные слои межсоединений, которые передают энергию и энергию. данные вокруг чипа.
Во-первых, говорит соучредитель Cyclos Мариос С. Папаефтимиу, профессор электротехники и компьютерных наук в Мичиганском университете, команда должна была найти способ включить спиральные петли катушек индуктивности в и без того довольно плотные слои межсоединений, которые передают энергию и энергию. данные вокруг чипа.
«Электросети устроены так, что в них полно петель, — говорит Папаефтимиу. «Дроссели расположены близко к линиям электропитания и к линиям заземления, и чем ближе они подходят к этим линиям, тем больше они им мешают». Команда нашла способ свести к минимуму помехи, сделав катушки индуктивности немного менее идеальными, намотав их по спирали, а также вверх и вниз через два верхних металлических слоя чипа. В схеме, разработанной командой для конструкции процессора AMD Piledriver, который составляет основу новых чипов Trinity компании, используется 9 процессоров.2 таких индуктора шириной 100 микрометров, распределенных по каждому модулю двухъядерного процессора. Основы разработки были представлены ранее в этом году на Международной конференции IEEE по твердотельным схемам.
Хотя тактовая мощность AMD снизилась на четверть, потенциальная экономия энергии может быть намного больше, говорит Мэтью Гутхаус, доцент компьютерной инженерии Калифорнийского университета в Санта-Круз. Его команда показала, что, настраивая размер катушек индуктивности и их расположение на кристалле, можно снизить тактовую мощность на целых 9 раз.0 процентов.
Наибольшая экономия достигается за счет уменьшения длины проводки между катушками индуктивности и триггерами в конце тактовой сети, которые неравномерно распределены по микросхеме. Команда Гутхауса представила такой подход к проектированию в июне на конференции по автоматизации проектирования в Сан-Франциско.
Несмотря на этот потенциал, до сих пор неясно, как далеко можно расширить резонансную синхронизацию. Не во всех микросхемах используются сетки часов, а катушки индуктивности могут занимать много места. Более того, хотя резонансные часы не обязательно должны работать непосредственно на резонансной частоте LC-контура, их эффективность как рециркулятора энергии снижается, когда часы работают значительно быстрее или медленнее. Это снижает полезность одной проверенной стратегии энергосбережения — временной работы чипов на более низкой скорости.
Это снижает полезность одной проверенной стратегии энергосбережения — временной работы чипов на более низкой скорости.
«Движение далеко от резонансной частоты не сэкономит энергию, а в какой-то момент [схема] вообще перестанет работать», — говорит Филлип Рестл, член исследовательского персонала Исследовательского центра Томаса Дж. Уотсона IBM в Йорктаун-Хайтс, штат Нью-Йорк. Компания Cyclos вставила переключатель, который позволяет чипу AMD включать и выключать резонансную часть часов. Но Рестл, который одним из первых работал над резонансными часами в микропроцессорах, говорит, что переключатель не является идеальным решением, потому что он увеличивает мощность, потребляемую чипом.
Тем не менее, Рестл говорит, что это не последний раз, когда мы увидим этот подход к тактированию. «Я считаю, что это первые дни для резонансных часов», — говорит он. «Есть еще кролики, которых нужно вытащить из шляпы».
Эта статья первоначально появилась в печати под названием «Хорошее время».