
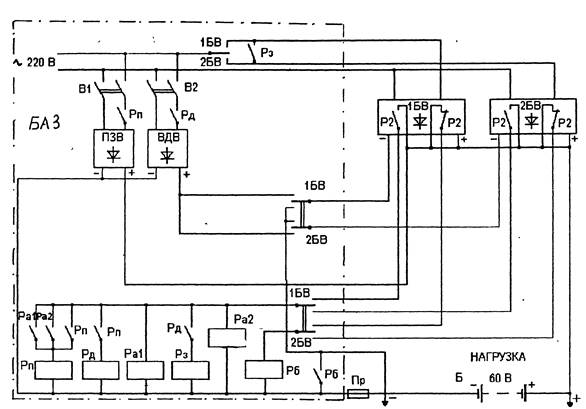
Электропитающая установка ЭПУ
Электропитающая установка ЭПУГлавнаяПродукцияСистемы бесперебойного питанияШкафы оперативного токаЭлектропитающая установка ЭПУ
Описание
Электропитающая установка (ЭПУ 220/48) предназначена для бесперебойного электроснабжение постоянным током напряжением 48 В, а также переменным током напряжением 220 В. Являестя источником бесперебойного питания т.м. модели SPE.
Отличительные особенности
Каждое ЭПУ разрабатывается и изготавливается строго ориентируясь на требования заказчика, это позволяет оптимизировать ЭПУ по таким параметрам как:
– функциональность;
– конструктив;
– эргономичность;
– возможность модернизации в процессе эксплуатации;
– стоимость.
Такие особенности ЭПУ позволяют легко интегрировать ее в систему энергоснабжения заказчика, а его эксплуатация существенно повышает надежность системы энергоснабжения, качество электроэнергии и уменьшает трудоемкость обслуживания системы электропитания.
Функции ЭПУ
— онлайн мониторинг и диагностика качество электроэнергии, состояние аппаратов защиты;
— формирование сигналов аварии при токах короткого замыкания, перенапряжения, выходе из строя узлов системы энергоснабжения;
— селективное отключение нагрузки;
— резервирование источник питания переменного и/или постоянного тока, в том числе двойное резервирование.
Сертификаты
ЭПУ соответствует требованиям ТР ТС 004/2011 и ТР ТС 020/201
Область применения ЭПУ
Потребители первой, особой категории, средства связи и телекоммуникаций, социально-значимые объекты.
Основные свойства и характеристика ЭПУ
|
Род тока вводной цепи |
Трехфазный/однофазный |
|
Номинальное напряжение на вводе AC , В |
380±15/220 |
|
Частота питающей сети, Гц |
50±5 |
|
Сопротивление изоляции |
свыше 20 МОм |
|
Степень защиты оболочки по ГОСТ 14254-96 |
IP 20 |
|
Номинальные токи автоматов отходящих линий, А |
в соответствии с таблицей экспликации |
|
Предельная отключающая способность при коротком замыкании, кА |
6 |
|
Вид системы заземления |
TN-С-S |
|
Вид разделения по ГОСТ 51321. |
1 |
|
Тип электрических соединений функциональных блоков |
FF |
|
Условия эксплуатации по ГОСТ 15150-69 |
УХЛ4 |
|
Номинальное переменное напряжение на выходе, В |
230±15 |
|
Номинальное постоянное напряжение на выходе, В |
48 |
|
Время работы от АКБ при 100% нагрузке, мин |
57 |
|
Установочные данные | |
|
Габаритные размеры ВхШхГ, мм не более |
2500х850х600 |
|
Тип конструктивного исполнения |
Шкафное |
|
Способ установки |
Напольный |
|
Подвод сети электроэнергии |
снизу шкафа |
|
Вывод отходящих линий |
|
 1-2000
1-2000Для чего нужны электропитающие установки, их особенности
05 ноября 2016
Электропитающие установки (ЭПУ) – электропитающая установка постоянного тока. Предназначена для надежного обеспечения электричеством промышленного оборудования разного типа назначения, номинальное напряжение постоянного тока составляет 24, 48, 60 В. Подобные установки распространены для резервного питания телекоммуникационной техники, автоматизированного оборудования, охранной и пожарной систем.
Предназначена для надежного обеспечения электричеством промышленного оборудования разного типа назначения, номинальное напряжение постоянного тока составляет 24, 48, 60 В. Подобные установки распространены для резервного питания телекоммуникационной техники, автоматизированного оборудования, охранной и пожарной систем.
Список основных электропитающих установок (ЭПУ):
- Преобразователь AC/DC;
- ВЧ преобразователь DC/DC;
- Система на аккумуляторах;
- Контроллер управления.
Существует ряд параметров, которые должны быть учтены при подборе установке электропитания. Среди них перечень важных функциональных и нормативных факторов.
Важные параметры:
- уровень мощности нагрузки, подключенной к ЭПУ;
- напряжение питания нагрузки;
- коэффициент одновременности нагрузки;
- требуемое время поддержки;
- параметры напряжения сети;
- время, максимально допустимое, для полной зарядки аккумулятора.

В отдельных случаях специальные нагрузки требуют большого пускового тока, в этом случае необходимо брать во внимание эти значения тока. Система должна быть надежной и гарантировать возможность производить мониторинг. В большей части наших систем реализуется мoдульная концепция параллельных установок, благодаря чему дополнительно увеличивается мощность, в том числе редундантность системы. Механика оборудования должна быть в 100% случаев быть готовой к полной установке модулей, благодаря чему система может быть в любой момент расширена с минимумом финансовых затрат.
Что необходимо знать об ЭПУ
В состав ЭПУ включены установки бесперебойного питания постоянного и переменного тока, преобразователи и стабилизаторы напряжения, коммутационное оборудование и токораспределительные сети, которые связывают между собой оборудование электропитания и аппаратуру связи. Важно, чтобы установка была рассчитана на работу в стандартном и аварийном режимах.
Если говорить о нормальном режиме работы, то он обеспечивает аппаратуре связи качество электроэнергии, которое соответствует установленным нормам, а сама ЭПУ должна работать без участия обслуживающего персонала. Переход в аварийный режим не обеспечит аппаратуре установленное качество электричества и потребует участия специалистов эксплуатации. Такой режим предполагает обеспечение автоматического отключения поврежденного оборудования и выдачу соответствующего сигнала.
Переход в аварийный режим не обеспечит аппаратуре установленное качество электричества и потребует участия специалистов эксплуатации. Такой режим предполагает обеспечение автоматического отключения поврежденного оборудования и выдачу соответствующего сигнала.
Оборудование может автоматически отключаться в случае если дальнейшая работа грозит необратимыми повреждениями оборудования, к примеру, разрядится аккумуляторная батарея ниже допустимого уровня, либо же если нарушаются требования безопасности.
Особенности установки
Можно произвести установку оборудования согласно индивидуального проекта. ЭПУ могут быть выполнен в соответствии с собственным проектом заказчика, диапазон единиц будет составлять до нескольких сотен киловатт. Некоторое оборудование имеет выходной ток до 3000 Ампер. В комплект должны быть включены удобные для подключения распределительные шкафы, которые оснащены всем, что требуется для включения необходимого количества потребителей со всеми требуемыми узлами контроля.
Загадка 1090 МГц
Рекомендовать правки
В ADS-B есть много параметров, которые определяют качество данных в отчетах о местоположении и скорости. Эти параметры служат индикаторами точности и достоверности информации о местоположении и скорости воздушного судна, передаваемой в сообщениях ADS-B.
С каждой эволюцией версий ADS-B эти параметры переименовывались и определялись. Эти обновления усложняют понимание и анализ неопределенностей в ADS-B. Эта глава предназначена для лучшего обзора индикаторов неопределенности в различных версиях ADS-B, а также дает четкое сопоставление между индикаторами и фактическими значениями неопределенности.
Терминология
В целом существует три типа индикаторов качества данных:
Индикаторы неопределенности допустимые пределы неопределенности.
Индикаторы точности : Эти индикаторы впервые представлены в версии 1 и предназначены для замены индикаторов неопределенности в версии 0.
Значения параметров также указывают, что не менее 95% измерений находятся в допустимых пределах точности.Индикаторы целостности : Эти индикаторы также впервые представлены в версии 1, заменив индикаторы неопределенности из версии 0.
 Значения параметров также указывают, что не менее 95% измерений находятся в допустимых пределах точности.
Значения параметров также указывают, что не менее 95% измерений находятся в допустимых пределах точности.Обычно два параметра связаны с положением и скоростью отдельно в каждой группе индикаторов. В таблице 1.1 показаны шесть основных показателей неопределенности, относящихся к версиям ADS-B, и диапазоны их значений.
| Категория навигационной неопределенности – положение | НУКП | 0 | 0–9 |
| Категория навигационной неопределенности – скорость (скорость) | НУКр | 0 | 0–4 |
| Категория точности навигации – позиция | НАКп | 1, 2 | 0–11 |
| Категория навигационной точности – скорость | НАКв | 1, 2 | 0–4 |
| Категория целостности навигации | Сетевая карта | 1, 2 | 0–11 |
| Уровень целостности наблюдения | Уровень безопасности | 1, 2 | 0–3 |
Каждый показатель качества также относится к набору параметров, которые выражают точное количество неопределенностей, ошибок или вероятностей.
|л|р8см|л| и Параметр и
NUCp (V0) и предел защиты по горизонтали и HPL
и радиус удержания 95 % при ошибке горизонтального положения и Rc/μ
и радиус удержания 95 % при ошибке вертикального положения и Rc/v
NUCr (V0) и ошибка горизонтальной скорости (95 %) и HVE
и ошибка вертикальной скорости (95 %) и VVE
NACp (V1,2) и расчетная неопределенность положения (горизонтальная точность 95 % — p). также известный как горизонтальный показатель качества (HFOM) в GNSS и EPU
и оценочная неопределенность положения по вертикали (95% точность по вертикали — п). также известный как вертикальный показатель качества (VFOM) в GNSS и VEPU
и вертикальный показатель качества (горизонтальная точность 95% — v) и VFOMr
NIC (V1,2) и ограничение горизонтального радиуса удержания и RC
и ограничение вертикальной защиты и VPL
SIL (V1,2) и вероятность превышения горизонтального радиуса удержания и P-RC
& вероятность превышения вертикальной целостности области удержания и P -ВПЛ
Эти параметры идентифицируются разными битами из разных сообщений. В связи с развитием версий ADS-B определения также могут различаться. Для правильного получения этих параметров необходимо определить версию ответчика ADS-B. Это может быть основано на логике, описанной в главе [chap:adsb-basic].
В связи с развитием версий ADS-B определения также могут различаться. Для правильного получения этих параметров необходимо определить версию ответчика ADS-B. Это может быть основано на логике, описанной в главе [chap:adsb-basic].
В оставшейся части этой главы эти параметры подробно объясняются в соответствии с различными версиями ADS-B. Также указаны изменения одних и тех же параметров между разными версиями.
Версия 0
В версии 0 ADS-B определяются только неопределенности. Два набора параметров связаны с положением и скоростью (или скоростью) отдельно.
Категория навигационной неопределенности – местоположение (NUCp)
NUCp напрямую связан с кодом типа ADS-B с однозначным отображением. Сопоставление кода типа и NUCp в сообщениях о местоположении на земле и двух типах сообщений о местоположении в воздухе показано в таблице 1.4.
0,7
| 0 | 5 | 6 | 7 | 8 | |
| НУКп | 0 | 9 | 8 | 7 | 6 |
0,7
| ТК | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 |
| НУКп | 9 | 8 | 7 | 6 | 5 | 4 | 3 | 2 | 1 | 0 |
0,7
| ТК | 20 | 21 | 22 |
| НУКп | 9 | 8 | 0 |
В целом, чем выше число NUCp (меньше число TC), тем выше достоверность измерения положения. При работе с неопределенностью положения для количественной оценки неопределенностей используются горизонтальный предел защиты (HPL), радиус локализации ошибки горизонтального положения (обозначается как Rc/μ) и радиус локализации ошибки вертикального положения (обозначается Rc/v). . Все значения приведены в таблице 1.5.
При работе с неопределенностью положения для количественной оценки неопределенностей используются горизонтальный предел защиты (HPL), радиус локализации ошибки горизонтального положения (обозначается как Rc/μ) и радиус локализации ошибки вертикального положения (обозначается Rc/v). . Все значения приведены в таблице 1.5.
| 0 | 0 | Н/Д | Н/Д | Н/Д |
| 5 | 9 | <7,5 м | <3 м | Н/Д |
| 6 | 8 | <25 м | <10 м | Н/Д |
| 7 | 7 | <0,1 м. мили (185 м) | <0,05 м. мили (93 м) | Н/Д |
| 8 | 6 | >0,1 м. мили (185 м) | >0,05 м. мили (93 м) | Н/Д |
| 9 | 9 | <7,5 м | <3 м | Н/Д |
| 10 | 8 | <25 м | <10 м | Н/Д |
| 11 | 7 | <0,1 м. мили (185 м) мили (185 м) | <0,05 м. мили (93 м) | Н/Д |
| 12 | 6 | <0,2 м. мили (370 м) | <0,1 м. мили (185 м) | Н/Д |
| 13 | 5 | <0,5 м. мили (926 м) | <0,25 м. мили (463 м) | Н/Д |
| 14 | 4 | <1 м. мили (1852 м) | <0,5 м. мили (926 м) | Н/Д |
| 15 | 3 | <2 морских миль (3704 м) | <1 м. мили (1852 м) | Н/Д |
| 16 | 2 | <10 морских миль (18520 м) | <5 морских миль (9260 м) | Н/Д |
| 17 | 1 | <20 морских миль (37040 м) | <10 морских миль (18520 м) | Н/Д |
| 18 | 0 | >20 морских миль (37040 м) | >10 морских миль (18520 м) | Н/Д |
| 20 | 9 | <7,5 м | <3 м | <4 м |
| 21 | 8 | <25 м | <10 м | <15 м |
| 22 | 0 | >25 м | >10 м | >15 м |
Стоит отметить, что в случае положения в воздухе с высотой по GNSS (TC=20–22) HPL и Rc/μ определяются с небольшими отличиями по сравнению с другими типами сообщений о местоположении. Кроме того, в этом сообщении определяется радиус удержания для вертикального положения (Rc/v). Это возможно, потому что высота получена из источников GNSS.
Кроме того, в этом сообщении определяется радиус удержания для вертикального положения (Rc/v). Это возможно, потому что высота получена из источников GNSS.
Категория навигационной неопределенности — скорость (NUCr)
Категория навигационной неопределенности — скорость (NUCr) используется для указания неопределенности горизонтальной и вертикальной скоростей. Биты, представляющие NUCr, можно найти в сообщении о скорости в воздухе (TC=19). NUCr расположен в битах сообщения 43–45 (или битах ME 11–13), что определяет 95% ошибки горизонтальной и вертикальной скорости, как показано в таблице 1.6.
| 0 | Н/Д | Н/Д |
| 1 | <10 м/с | <15,2 м/с (50 кадр/с) |
| 2 | <3 м/с | <4,5 м/с (15 кадр/с) |
| 3 | <1 м/с | <1,5 м/с (5 кадр/с) |
| 4 | <0,3 м/с | <0,46 м/с (1,5 кадр/с) |
Версия 1
В версии 1 ADS-B категория неопределенности удалена и заменена категорией точности и категорией целостности. И NUCp, и NUCr из версии 0 больше не существуют в версии 1. Новые индикаторы, представленные в версии 1, — это NACp, NACv, NIC и SIL.
И NUCp, и NUCr из версии 0 больше не существуют в версии 1. Новые индикаторы, представленные в версии 1, — это NACp, NACv, NIC и SIL.
Категория целостности навигации (NIC)
NIC предназначен для замены NUCp версии 0, но включает больше уровней. Как и NUCp в версии 0, категория целостности навигации (NIC) связана с кодом типа. Однако Type Code и NIC больше не имеют взаимно однозначного сопоставления. При определении большего количества уровней требуется дополнительный бит, чтобы различать два уровня, представленных некоторыми одинаковыми кодами типов.
Бит дополнения NIC (NICs) вводится в сообщениях о рабочем состоянии (TC=31) и располагается в бите 76 сообщения (или бите 44 ME). Отношения между TC, NIC и Rc перечислены в таблице 1.7.
| 0 | Н/Д | Н/Д | Н/Д | Н/Д |
| 5 | 0 | 11 | <7,5 м | Н/Д |
| 6 | 0 | 10 | <25 м | Н/Д |
| 7 | 1 | 9 | <75 м | Н/Д |
| 0 | 8 | <0,1 м. мили (185 м) мили (185 м) | Н/Д | |
| 8 | 0 | 0 | >0,1 м. мили или неизвестно | Н/Д |
| 9 | 0 | 11 | <7,5 м | <11 м |
| 10 | 0 | 10 | <25 м | <37,5 м |
| 11 | 1 | 9 | <75 м | <112 м |
| 0 | 8 | <0,1 м. мили (185 м) | Н/Д | |
| 12 | 0 | 7 | <0,2 м. мили (370 м) | Н/Д |
| 13 | 0 | 6 | <0,5 м. мили (926 м) | Н/Д |
| 1 | <0,6 м. мили (1111 м) | Н/Д | ||
| 14 | 0 | 5 | <1,0 м. мили (1852 м) | Н/Д |
| 15 | 0 | 4 | <2 морских миль (3704 м) | Н/Д |
| 16 | 1 | 3 | <4 морских миль (7408 м) | Н/Д |
| 0 | 2 | <8 морских миль (14,8 км) | Н/Д | |
| 17 | 0 | 1 | <20 морских миль (37,0 км) | Н/Д |
| 18 | 0 | 0 | >20 морских миль или неизвестно | Н/Д |
| 20 | 0 | 11 | <7,5 м | <11 м |
| 21 | 0 | 10 | <25 м | <37,5 м |
| 22 | 0 | 0 | >25 м | >112 м |
Категория навигационной точности – положение (NACp)
NACp введен в ADS-B версии 1 в качестве дополнительного индикатора NIC. NACp можно получить из сообщения о рабочем состоянии, биты 77–80 кадра сообщения (или биты 45–48 ME).
NACp можно получить из сообщения о рабочем состоянии, биты 77–80 кадра сообщения (или биты 45–48 ME).
С помощью NACp можно определить границы горизонтальной и вертикальной точности 95 %. Это оценочная неопределенность положения (EPU) и оценочная неопределенность положения по вертикали (VEPU), которые также известны как показатель качества по горизонтали (HFOM) и показатель качества по вертикали (VFOM) в GNSS.
EPU и Rc в NIC имеют следующую связь:
\[\begin{split} \mathrm{EPU} &\приблизительно \mathrm{Rc} / 2.5 \qquad \text{для NACp} \ge 9 \\ \mathrm{EPU} &\приблизительно \mathrm{Rc} / 2.0 \qquad \text{для NACp} < 9 \end{split}\]
NACp и значения связанных с ним параметров определены в таблице 1.8.
| 11 | <3 м | <4 м |
| 10 | <10 м | <15 м |
| 9 | <30 м | <45 м |
| 8 | <0,05 м. мили (93 м) мили (93 м) | Н/Д |
| 7 | <0,1 м. мили (185 м) | Н/Д |
| 6 | <0,3 м. мили (556 м) | Н/Д |
| 5 | <0,5 м. мили (926 м) | Н/Д |
| 4 | <1,0 м. мили (1852 м) | Н/Д |
| 3 | <2 морских миль (3704 м) | Н/Д |
| 2 | <4 морских миль (7408 м) | Н/Д |
| 1 | <10 морских миль (18520 м) | Н/Д |
| 0 | >10 морских миль или неизвестно | Н/Д |
Категория точности навигации – скорость (NACv)
NACv введен в версии 1 для замены NUCv версии 0. Биты расположены в том же месте и имеют те же определения значений. Эти биты содержатся в сообщении о скорости в воздухе (TC=19).) биты сообщения 43–45 (или биты ME 11–13). Он определяет 95% ошибок горизонтальной и вертикальной скоростей. Подробные определения Горизонтального показателя качества для ставки (HFOMr) и Вертикального показателя качества для ставки (HFOMr) приведены в Таблице 1.9.
Подробные определения Горизонтального показателя качества для ставки (HFOMr) и Вертикального показателя качества для ставки (HFOMr) приведены в Таблице 1.9.
| 0 | Н/Д | Н/Д |
| 1 | <10 м/с | <15,2 м/с (50 кадр/с) |
| 2 | <3 м/с | <4,5 м/с (15 кадр/с) |
| 3 | <1 м/с | <1,5 м/с (5 кадр/с) |
| 4 | <0,3 м/с | <0,46 м/с (1,5 кадр/с) |
Уровень целостности наблюдения (SIL)
SIL введен в версии 1 и используется для указания вероятности измерений, превышающих радиус локализации. Значение SIL также можно найти в сообщении о рабочем состоянии (TC=31), биты сообщения 83–84 (или биты ME 51–52) 9{-7}\)
Единицей измерения для P-RCu и P-VPL может быть летный час или образец, за исключением случаев, когда SIL=3, единицей измерения для P-VPL становятся 150 секунд на образец.
Версия 2
Между версией 1 и версией 2 меньше изменений по сравнению с предыдущим обновлением. Основными изменениями являются дальнейшие уточнения уровней NIC и незначительные обновления параметров SIL.
Категория целостности навигации (NIC)
В версии 2 уровни NIC можно получить с помощью трех дополнительных дополнительных битов, называемых дополнительным битом NIC A (NICa), дополнительным битом NIC B (NICb) и дополнительным битом NIC C (NICc). Эти биты можно найти следующим образом:
NICa находится в сообщении о рабочем состоянии (TC=31) (76-й бит сообщения или 44-й бит ME), что совпадает с ADS-B версии 1.
NICb находится в сообщении о местоположении в воздухе (TC=9–18), бит 40 сообщения или бит 8 ME), где в предыдущих версиях ADS-B располагался флаг одиночной антенны.
NICc также содержится в сообщении о рабочем состоянии (TC=31) (бит 52 сообщения или бит 20 ME).
С помощью этих дополнительных битов и кода типа можно рассчитать значение NIC в версии 2. Значения NIC и соответствующие им значения Rc показаны в таблице 1.11.
Значения NIC и соответствующие им значения Rc показаны в таблице 1.11.
| 5 | 0 | Н/Д | 0 | 11 | <7,5 м |
| 6 | 0 | Н/Д | 0 | 10 | <25 м |
| 7 | 1 | Н/Д | 0 | 9 | <75 м |
| 0 | Н/Д | 0 | 8 | <0,1 м. мили (185 м) | |
| 8 | 1 | Н/Д | 1 | 7 | <0,2 м. мили (370 м) |
| 1 | Н/Д | 0 | 6 | <0,3 м. мили (556 м) | |
| 0 | Н/Д | 1 | <0,6 м. мили (1111 м) | ||
| 0 | Н/Д | 0 | 0 | >0,6 м. мили или неизвестно | |
| 9 | 0 | 0 | Н/Д | 11 | <7,5 м |
| 10 | 0 | 0 | Н/Д | 10 | <25 м |
| 11 | 1 | 1 | Н/Д | 9 | <75 м |
| 0 | 0 | Н/Д | 8 | <0,1 м. мили (185 м) мили (185 м) | |
| 12 | 0 | 0 | Н/Д | 7 | <0,2 м. мили (370 м) |
| 13 | 0 | 1 | Н/Д | 6 | <0,3 м. мили (556 м) |
| 0 | 0 | Н/Д | <0,5 м. мили (926 м) | ||
| 1 | 1 | Н/Д | <0,6 м. мили (1111 м) | ||
| 14 | 0 | 0 | Н/Д | 5 | <1,0 м. мили (1852 м) |
| 15 | 0 | 0 | Н/Д | 4 | <2 морских миль (3704 м) |
| 16 | 1 | 1 | Н/Д | 3 | <4 морских миль (7408 м) |
| 0 | 0 | Н/Д | 2 | <8 морских миль (14,8 км) | |
| 17 | 0 | 0 | Н/Д | 1 | <20 морских миль (37,0 км) |
| 18 | 0 | 0 | Н/Д | 0 | >20 морских миль или неизвестно |
| 20 | Н/Д | Н/Д | Н/Д | 11 | <7,5 м |
| 21 | Н/Д | Н/Д | Н/Д | 10 | <25 м |
| 22 | Н/Д | Н/Д | Н/Д | 0 | >25 м |
Уровень полноты наблюдения (SIL)
В версии 2 введен дополнительный дополнительный бит SIL (SIL), чтобы лучше различать, имеет ли значение единицу измерения: летный час или выборка. Бит SILs также можно найти в сообщении о рабочем состоянии, бит 87 сообщения (или бит 55 ME). Определения:
Бит SILs также можно найти в сообщении о рабочем состоянии, бит 87 сообщения (или бит 55 ME). Определения:
Значения, относящиеся к SIL, остаются такими же, как показано в таблице 1.10.
NACp и NACv
NACp и NACv в версии 2 остаются такими же, как и в версии 1. Соответствующие параметры и определения можно найти в предыдущих таблицах 1.8 и 1.9 соответственно.
Расшифровка состояний мРНК рибосомы млекопитающих с помощью трансляционных комплексов ГТФазы
Адамс П.Д., Афонин П.В., Бункоци Г., Чен В.Б., Дэвис И.В., Эколс Н., Хедд Дж.Дж., Хунг Л.-В., Капрал Г.Дж., Гроссе-Кунстлеве RW PHENIX: комплексная система на основе Python для решения макромолекулярной структуры. Акта Кристаллогр. Д биол. Кристаллогр. 2010;66:213–221. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Алксне Л.Э., Энтони Р.А., Либман С.В., Уорнер Дж.Р. Центр точности в рибосоме сохраняется более 2 миллиардов лет. проц. Натл. акад. науч. США. 1993;90:9538–9541. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Амунтс А. , Браун А., Бай X.-C., Лласер Дж.Л., Хуссейн Т., Эмсли П., Лонг Ф., Муршудов Г., Шерес С.Х.В. , Рамакришнан В. Структура большой рибосомной субъединицы митохондрий дрожжей. Наука. 2014; 343:1485–1489. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Браун А., Бай X.-C., Лласер Дж.Л., Хуссейн Т., Эмсли П., Лонг Ф., Муршудов Г., Шерес С.Х.В. , Рамакришнан В. Структура большой рибосомной субъединицы митохондрий дрожжей. Наука. 2014; 343:1485–1489. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Андерсен Г.Р., Педерсен Л., Валенте Л., Чаттерджи И., Кинзи Т.Г., Кьельдгаард М., Ниборг Дж. Структурная основа обмена нуклеотидов и конкуренции с тРНК в комплексе факторов элонгации дрожжей eEF1A:eEF1Balpha. Мол. Клетка. 2000; 6: 1261–1266. [PubMed] [Google Scholar]
Andersson D.I., van Verseveld H.W., Stouthamer A.H., Kurland C.G. Субоптимальный рост со сверхточными рибосомами. Арка микробиол. 1986; 144: 96–101. [PubMed] [Google Scholar]
Аткинсон Дж. К., Бальдауф С. Л., Хаурилюк В. Эволюция безостановочного, бездействующего и нонсенс-опосредованного распада мРНК и их компонентов, производных от факторов терминации. БМС Эвол. биол. 2008;8:290. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Bai X.-C., Rajendra E.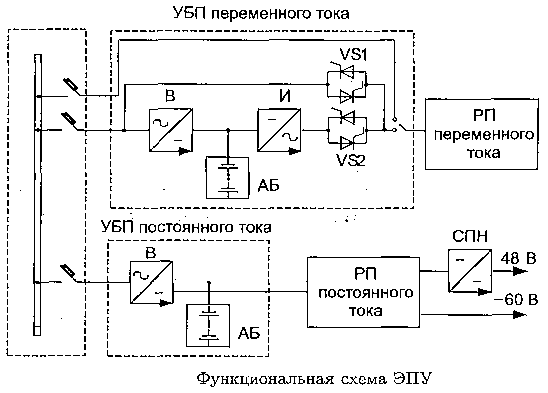 , Yang G., Shi Y., Scheres S.H. Выборка конформационного пространства каталитической субъединицы γ-секретазы человека. электронная жизнь. 2015;4:e11182. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Yang G., Shi Y., Scheres S.H. Выборка конформационного пространства каталитической субъединицы γ-секретазы человека. электронная жизнь. 2015;4:e11182. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Becker T., Armache J.-P., Jarasch A., Anger A.M., Villa E., Sieber H., Motaal B.A., Mielke T., Berninghausen O. , Бекманн Р. Структура комплекса распада мРНК Dom34-Hbs1, связанного с застопорившейся рибосомой 80S. Нац. Структура Мол. биол. 2011;18:715–720. [PubMed] [Академия Google]
Беккер Т., Франкенберг С., Виклс С., Шумейкер С.Дж., Ангер А.М., Армаш Дж.-П., Зибер Х., Унгевикелл С., Бернингхаузен О., Даберков И. Структурные основы высококонсервативной рециркуляции рибосом в эукариоты и археи. Природа. 2012; 482: 501–506. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Бишофф Л., Бернингхаузен О., Бекманн Р. Молекулярная основа рибосомы, функционирующей как датчик L-триптофана. Cell Rep. 2014; 9: 469–475. [PubMed] [Google Scholar]
Blanchard SC, Gonzalez RL, Kim HD, Chu S. , Puglisi JD Выбор тРНК и кинетическая корректура в переводе. Нац. Структура Мол. биол. 2004; 11:1008–1014. [PubMed] [Академия Google]
, Puglisi JD Выбор тРНК и кинетическая корректура в переводе. Нац. Структура Мол. биол. 2004; 11:1008–1014. [PubMed] [Академия Google]
Браун А., Лонг Ф., Николлс Р.А., Тутс Дж., Эмсли П., Муршудов Г. Инструменты для построения макромолекулярных моделей и уточнения реконструкций электронной криомикроскопии. Акта Кристаллогр. Д биол. Кристаллогр. 2015;71:136–153. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Браун А., Шао С., Мюррей Дж., Хегде Р.С., Рамакришнан В. Структурная основа распознавания стоп-кодонов у эукариот. Природа. 2015; 524:493–496. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Бруно И.Дж., Коул Дж.К., Кесслер М., Луо Дж., Мазервелл В.Д., Пуркис Л.Х., Смит Б.Р., Тейлор Р., Купер Р.И., Харрис С.Е., Орпен А.Г. кристаллографически полученной информации о молекулярной геометрии. J. Chem. Инф. вычисл. науч. 2004;44:2133–2144. [PubMed] [Академия Google]
Карелли Дж.Д., Сетофер С.Г., Смит Г.А., Миллер Х.Р., Симард Дж.Л., Меррик В.К., Джейн Р. К., Росс Н.Т., Тонтон Дж. Тернатин и его улучшенные синтетические варианты убивают раковые клетки, воздействуя на тройной комплекс фактор элонгации-1А. электронная жизнь. 2015;4:e10222. [Бесплатная статья PMC] [PubMed] [Google Scholar]
К., Росс Н.Т., Тонтон Дж. Тернатин и его улучшенные синтетические варианты убивают раковые клетки, воздействуя на тройной комплекс фактор элонгации-1А. электронная жизнь. 2015;4:e10222. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Chauvin C., Salhi S., Le Goff C., Viranaicken W., Diop D., Jean-Jean O. Участие человеческих факторов высвобождения eRF3a и eRF3b в переводе терминация и регуляция образования комплекса терминации. Мол. Клетка. биол. 2005; 25: 5801–5811. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Чен В.Б., Арендал В.Б., 3-й, Хедд Дж.Дж., Киди Д.А., Иммормино Р.М., Капрал Г.Дж., Мюррей Л.В., Ричардсон Дж.С., Ричардсон Д.К. MolProbity: проверка структуры всех атомов для макромолекулярной кристаллографии. Акта Кристаллогр. Д биол. Кристаллогр. 2010;66:12–21. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Чен С., Макмаллан Г., Фаруки А.Р., Муршудов Г.Н., Шорт Дж.М., Шерес С.Х.В., Хендерсон Р. Замена шума высокого разрешения для измерения переоснащения и проверки разрешения в 3D определение структуры с помощью электронной криомикроскопии отдельных частиц. Ультрамикроскопия. 2013; 135:24–35. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Ультрамикроскопия. 2013; 135:24–35. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Ченг З., Сайто К., Писарев А.В., Вада М., Писарева В.П., Пестова Т.В., Гайда М., Раунд А., Конг С., Лим М. Структурное понимание eRF3 и распознавание стоп-кодона eRF1. Гены Дев. 2009; 23:1106–1118. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Cochella L., Green R. Активная роль тРНК в декодировании за пределами спаривания кодон:антикодон. Наука. 2005; 308:1178–1180. [PMC free article] [PubMed] [Google Scholar]
Крепин Т., Шалак В.Ф., Яремчук А.Д., Власенко Д.О., Маккарти А., Негруцкий Б.С., Тукало М.А., Ельская А.В. Фактор элонгации трансляции eEF1A2 млекопитающих: рентгеновская структура и новые особенности механизма обмена GDP/GTP у высших эукариот. Нуклеиновые Кислоты Res. 2014;42:12939–12948. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Crews C.M., Collins J.L., Lane W.S., Snapper M.L., Schreiber S.L. ГТФ-зависимое связывание антипролиферативного агента дидемнина с фактором элонгации 1 альфа. Дж. Биол. хим. 1994; 269:15411–15414. [PubMed] [Google Scholar]
Дж. Биол. хим. 1994; 269:15411–15414. [PubMed] [Google Scholar]
Демешкина Н., Дженнер Л., Вестхоф Э., Юсупов М., Юсупова Г. Новое понимание принципа декодирования на рибосоме. Природа. 2012; 484: 256–259. [PubMed] [Google Scholar]
Девер Т.Э., Грин Р. Фазы удлинения, терминации и рециркуляции трансляции у эукариот. Харб Колд Спринг. Перспектива. биол. 2012;4:a013706. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Эмсли П., Локамп Б., Скотт В.Г., Коутан К. Особенности и развитие Coot. Акта Кристаллогр. Д биол. Кристаллогр. 2010;66:486–501. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Faxén M., Walles-Granberg A., Isaksson L.A. Антисупрессия за счет мутации в rpsM(S13), дающей укороченный рибосомный белок S13. Биохим. Биофиз. Акта. 1994;1218:27–34. [PubMed] [Google Scholar]
Фишер Н., Нейманн П., Коневега А.Л., Бок Л.В., Фикнер Р., Роднина М.В., Старк Х. Структура комплекса рибосома-EF-Tu E. coli при разрешении <3 Å крио-ЭМ с поправкой на Cs. Природа. 2015; 520: 567–570. [PubMed] [Академия Google]
Природа. 2015; 520: 567–570. [PubMed] [Академия Google]
Гейгер Т., Венер А., Шааб К., Кокс Дж., Манн М. Сравнительный протеомный анализ одиннадцати распространенных клеточных линий показывает повсеместную, но различную экспрессию большинства белков. Мол. Клеточная протеомика. 2012;11 М111.014050. [Бесплатная статья PMC] [PubMed] [Google Scholar]
des Georges A., Hashem Y., Unbehaun A., Grassucci R.A., Taylor D., Hellen C.U.T., Pestova T.V., Frank J. Структура рибосомных пре- комплекс терминации, связанный с eRF1 ∗ eRF3 ∗ GDPNP. Нуклеиновые Кислоты Res. 2014;42:3409–3418. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Гайдош Н.Р., Грин Р. Dom34 спасает рибосомы в 3′-нетранслируемых областях. Клетка. 2014; 156:950–962. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Хоссейн М.Б., ван дер Хельм Д., Антел Дж., Шелдрик Г.М., Сандуя С.К., Вайнхаймер А.Дж. Кристаллическая и молекулярная структура дидемнина В, противовирусного и цитотоксического депсипептида. проц. Натл. акад. науч. США. 1988; 85: 4118–4122. [PMC free article] [PubMed] [Google Scholar]
проц. Натл. акад. науч. США. 1988; 85: 4118–4122. [PMC free article] [PubMed] [Google Scholar]
Дженнер Л.Б., Демешкина Н., Юсупова Г., Юсупов М. Структурные аспекты поддержания рамки считывания матричной РНК рибосомой. Нац. Структура Мол. биол. 2010;17:555–560. [PubMed] [Академия Google]
Клинк Б.У., Гуди Р.С., Шайдиг А.Дж. Недавно разработанный микроспектрофлуориметр для кинетических исследований белковых кристаллов в сочетании с рентгеновской дифракцией. Биофиз. Дж. 2006; 91:981–992. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Кобаяси К., Кикуно И., Куроха К., Сайто К., Ито К., Ишитани Р., Инада Т., Нуреки О. Структурная основа для наблюдения за мРНК архейной Pelota и комплексом EF1α, связанным с GTP. проц. Натл. акад. науч. США. 2010;107:17575–17579. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Кобаяши К., Сайто К., Ишитани Р., Ито К., Нуреки О. Структурная основа терминации трансляции архейным RF1 и комплексом EF1α, связанным с ГТФ. Нуклеиновые Кислоты Res. 2012;40:9319–9328. [Бесплатная статья PMC] [PubMed] [Google Scholar]
2012;40:9319–9328. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Крамер Э. Б., Валлабханени Х., Майер Л. М., Фарабо П. Дж. Всесторонний анализ трансляционных миссенс-ошибок у дрожжей Saccharomyces cerevisiae. РНК. 2010;16:1797–1808. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Крастел П., Рогго С., Ширле М., Росс Н.Т., Перруччо Ф., Аспези П. мл., Ост Т., Бантин К., Эстоппи Д. ., Liechty B. Нанноцистин A: ингибитор фактора элонгации 1 из миксобактерий с различными противораковыми свойствами. Ангью. хим. Междунар. Эд. англ. 2015;54:10149–10154. [PubMed] [Google Scholar]
Кукукельбир А., Сигуорт Ф.Дж., Тагаре Х.Д. Количественная оценка локального разрешения крио-ЭМ карт плотности. Нац. Методы. 2014; 11:63–65. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Lee H.H., Kim Y.-S., Kim K.H., Heo I., Kim S.K., Kim O., Kim H.K., Yoon J.Y., Kim H.S., Kim D.J. Структурное и функциональное понимание Dom34, ключевого компонента нераспада мРНК. Мол. Клетка. 2007; 27: 938–950. [PubMed] [Google Scholar]
2007; 27: 938–950. [PubMed] [Google Scholar]
Li L.H., Timmins L.G., Wallace T.L., Krueger W.C., Prairie MD, Im W.B. Механизм действия дидемнина В, депсипептида из моря. Рак Летт. 1984;23:279–288. [PubMed] [Google Scholar]
Li X., Mooney P., Zheng S., Booth C.R., Braunfeld M.B., Gubbens S., Agard D.A., Cheng Y. одночастичная крио-ЭМ с разрешением. Нац. Методы. 2013; 10: 584–590. [PMC free article] [PubMed] [Google Scholar]
Loenarz C., Sekirnik R., Thalhammer A., Ge W., Spivakovsky E., Mackeen M.M., McDonough M.A., Cockman M.E., Kessler B.M., Ratcliffe P.J. эукариотический рибосомный центр декодирования влияет на точность трансляции. проц. Натл. акад. науч. США. 2014;111:4019–4024. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Матейсл С., Бернингхаузен О., Беккер Т., Бекманн Р. Структура комплекса терминации человеческого перевода. Нуклеиновые Кислоты Res. 2015;43:8615–8626. [Бесплатная статья PMC] [PubMed] [Google Scholar]
McCaughan KK, Brown C. M., Dalphin M.E., Berry MJ, Tate W.P. На эффективность терминации трансляции у млекопитающих влияет основание, следующее за стоп-кодоном. проц. Натл. акад. науч. США. 1995; 92: 5431–5435. [Бесплатная статья PMC] [PubMed] [Google Scholar]
M., Dalphin M.E., Berry MJ, Tate W.P. На эффективность терминации трансляции у млекопитающих влияет основание, следующее за стоп-кодоном. проц. Натл. акад. науч. США. 1995; 92: 5431–5435. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Мухс М., Хилал Т., Мильке Т., Скабкин М.А., Санбонмацу К.Ю., Пестова Т.В., Шпан С.М.Т. Крио-ЭМ рибосомных комплексов 80S с факторами терминации выявляет транслоцированный вирус паралича сверчков IRES. Мол. Клетка. 2015; 57: 422–432. [PMC free article] [PubMed] [Google Scholar]
Муршудов Г.Н., Скубак П., Лебедев А.А., Панну Н.С., Штайнер Р.А., Николлс Р.А., Винн М.Д., Лонг Ф., Вагин А.А. REFMAC5 для уточнения макромолекулярных кристаллических структур. Акта Кристаллогр. Д биол. Кристаллогр. 2011;67:355–367. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Ноеске Дж., Вассерман М.Р., Терри Д.С., Альтман Р.Б., Бланшар С.К., Кейт Дж.Х.Д. Структура рибосомы Escherichia coli с высоким разрешением. Нац. Структура Мол. биол. 2015;22:336–341. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
Огл Дж. М., Рамакришнан В. Структурное понимание точности перевода. Анну. Преподобный Биохим. 2005; 74: 129–177. [PubMed] [Google Scholar]
Огл Дж. М., Бродерсен Д. Э., Клемонс В. М., младший, Тарри М. Дж., Картер А. П., Рамакришнан В. Распознавание родственной транспортной РНК 30S рибосомной субъединицей. Наука. 2001;292: 897–902. [PubMed] [Google Scholar]
Огл Дж. М., Мерфи Ф. В., Тарри М. Дж., Рамакришнан В. Отбор тРНК рибосомой требует перехода от открытой формы к закрытой. Клетка. 2002; 111: 721–732. [PubMed] [Google Scholar]
Папе Т., Винтермейер В., Роднина М.В. Полный кинетический механизм зависимого от фактора элонгации связывания аминоацил-тРНК с сайтом A рибосомы E. coli. EMBO J. 1998; 17: 7490–7497. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Parmeggiani A., Krab I.M., Okamura S., Nielsen RC, Nyborg J., Nissen P. Структурная основа действия пульвомицина и GE2270 A на фактор удлинения Tu. Биохимия. 2006; 45: 6846–6857. [PubMed] [Академия Google]
Биохимия. 2006; 45: 6846–6857. [PubMed] [Академия Google]
Паскуалато С., Черфилс Дж. Кристаллографические доказательства гидролиза ГТФ с помощью субстрата небольшим белком, связывающим ГТФ. Состав. 2005; 13: 533–540. [PubMed] [Google Scholar]
Петтерсен Э.Ф., Годдард Т.Д., Хуанг К.С., Коуч Г.С., Гринблатт Д.М., Мэн Э.К., Феррин Т.Е. UCSF Chimera — система визуализации для поисковых исследований и анализа. Дж. Вычисл. хим. 2004; 25:1605–1612. [PubMed] [Google Scholar]
Пипенбург О., Папе Т., Плейсс Я.А., Винтермейер В., Уленбек О.К., Роднина М.В. Интактная аминоацил-тРНК необходима для запуска гидролиза GTP под действием фактора элонгации Tu на рибосоме. Биохимия. 2000;39: 1734–1738. [PubMed] [Google Scholar]
Писарева В.П., Скабкин М.А., Хеллен Ц.У.Т., Пестова Т.В., Писарев А.В. Диссоциация с помощью Pelota, Hbs1 и ABCE1 вакантных 80S рибосом млекопитающих и комплексов остановленной элонгации. EMBO J. 2011; 30: 1804–1817. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Preis A. , Heuer A., Barrio-Garcia C., Hauser A., Eyler D.E., Berninghausen O., Green R., Becker T., Beckmann R. Криоэлектронно-микроскопические структуры эукариотических комплексов терминации трансляции, содержащих eRF1-eRF3 или eRF1-ABCE1. Представитель ячейки 2014; 8:59–65. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Heuer A., Barrio-Garcia C., Hauser A., Eyler D.E., Berninghausen O., Green R., Becker T., Beckmann R. Криоэлектронно-микроскопические структуры эукариотических комплексов терминации трансляции, содержащих eRF1-eRF3 или eRF1-ABCE1. Представитель ячейки 2014; 8:59–65. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Rinehart K.L., Jr., Gloer J.B., Hughes R.G., Jr., Renis H.E., McGovren J.P., Swynenberg E.B., Stringfellow D.A., Kuentzel S.L., Li L.H. Didemnins: Antiviral and противоопухолевые депсипептиды из карибской оболочника. Наука. 1981; 212: 933–935. [PubMed] [Google Scholar]
Розенталь П.Б., Хендерсон Р. Оптимальное определение ориентации частиц, абсолютной руки и потери контраста в одночастичной электронной криомикроскопии. Дж. Мол. биол. 2003; 333: 721–745. [PubMed] [Академия Google]
Сайто К., Кобаяши К., Вада М., Кикуно И., Такусагава А., Мочизуки М., Утиуми Т., Иситани Р., Нуреки О., Ито К. Всемогущая роль фактора удлинения архей 1 альфа ( EF1α в трансляционной элонгации и терминации, а также в контроле качества синтеза белка, Proc. Natl. Acad. Sci. USA, 2010; 107: 19242–19247. коррекция индуцированного движения для субмегадальтонных крио-ЭМ частиц. eLife. 2014;3:e03665. [бесплатная статья PMC] [PubMed] [Google Scholar]
Natl. Acad. Sci. USA, 2010; 107: 19242–19247. коррекция индуцированного движения для субмегадальтонных крио-ЭМ частиц. eLife. 2014;3:e03665. [бесплатная статья PMC] [PubMed] [Google Scholar]
Шерес Ш.Х.В. Полуавтоматическая селекция крио-ЭМ частиц в РЕЛИОН-1.3. J. Структура. биол. 2015; 189:114–122. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Schmeing TM, Voorhees RM, Kelley AC, Gao Y.-GG, Murphy FV, 4th, Weir JR, Ramakrishnan V. Кристаллическая структура рибосомы, связанной с EF- Ту и аминоацил-тРНК. Наука. 2009; 326: 688–694. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Шао С., фон дер Мальсбург К., Хегде Р.С. Листерин-зависимое убиквитинирование зарождающегося белка зависит от диссоциации субъединиц рибосомы. Мол. Клетка. 2013;50:637–648. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Шарма А., Мариаппан М., Аппатурай С., Хегде Р.С. Рассечение транслокации белка в эндоплазматический ретикулум млекопитающих in vitro. Методы Мол. биол. 2010;619:339–363. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
Шумейкер С.Дж., Грин Р. Кинетический анализ показывает упорядоченное сочетание терминации трансляции и рециркуляции рибосом у дрожжей. проц. Натл. акад. науч. США. 2011;108:E1392–E1398. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Shoemaker CJ, Green R. Перевод обеспечивает контроль качества мРНК. Нац. Структура Мол. биол. 2012;19: 594–601. [Статья бесплатно PMC] [PubMed] [Google Scholar]
Шумейкер С.Дж., Эйлер Д.Е., Грин Р. Dom34:Hbs1 способствует диссоциации субъединиц и выпадению пептидил-тРНК, чтобы инициировать непрогрессирующий распад. Наука. 2010;330:369–372. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Tang G., Peng L., Baldwin P.R., Mann D.S., Jiang W., Rees I., Ludtke S.J., EMAN2 EMAN2: расширяемый пакет обработки изображений для электронной микроскопии . J. Структура. биол. 2007; 157:38–46. [PubMed] [Академия Google]
Тейлор Д., Унбехаун А., Ли В., Дас С., Лей Дж., Ляо Х.Ю. , Грассуччи Р.А., Пестова Т.В., Франк Дж. Крио-ЭМ структура комплекса терминации, связанного с фактором высвобождения эукариот eRF1-eRF3 у млекопитающих . проц. Натл. акад. науч. США. 2012;109:18413–18418. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Грассуччи Р.А., Пестова Т.В., Франк Дж. Крио-ЭМ структура комплекса терминации, связанного с фактором высвобождения эукариот eRF1-eRF3 у млекопитающих . проц. Натл. акад. науч. США. 2012;109:18413–18418. [Бесплатная статья PMC] [PubMed] [Google Scholar]
van den Elzen A.M.G., Henri J., Lazar N., Gas ME, Durand D., Lacroute F., Nicaise M., van Tilbeurgh H., Séraphin B. , Graille M. Вскрытие Dom34-Hbs1 выявило независимые функции в двух путях контроля качества РНК. Нац. Структура Мол. биол. 2010;17:1446–1452. [PubMed] [Академия Google]
Веттер И.Р., Виттингхофер А. Переключатель связывания гуаниновых нуклеотидов в трех измерениях. Наука. 2001; 294:1299–1304. [PubMed] [Google Scholar]
Вурхис Р.М., Рамакришнан В. Структурные основы цикла трансляционной элонгации. Анну. Преподобный Биохим. 2013; 82: 203–236. [PubMed] [Google Scholar]
Voorhees RM, Schmeing TM, Kelley AC, Ramakrishnan V. Механизм активации гидролиза GTP на рибосоме. Наука. 2010; 330:835–838.