
Особенности экспрессии молекул-маркеров инсулинорезистентности при экспериментальной болезни Альцгеймера
По мере старения населения число страдающих деменцией людей, по прогнозам ученых, к 2050 г. возрастет до 115 млн, что, несомненно, создаст значимую проблему для здравоохранения [1]. Клинически заболевание проявляется прогрессирующей потерей памяти и постепенным снижением когнитивных функций [2]. Болезнь Альцгеймера (БА) характеризуется значительной потере нейронов и синапсов, особенно в гиппокампе и коре больших полушарий, внеклеточным накоплением β-амилоида и формированием нейрофибриллярных сплетений [3]. Однако диагноз устанавливается, как правило, на стадии значительного повреждения головного мозга, что существенно снижает эффективность лечения [4]. Большое значение может иметь идентификация молекул-маркеров, позволяющая выявить заболевание на ранних стадиях у людей с высоким риском развития слабоумия.
Результаты недавних исследований свидетельствуют о ключевой роли инсулинорезистентности в развитии когнитивных нарушений и нейродегенерации, в частности при БА [5, 6].
Нарушение передачи сигналов инсулина в головном мозге инициирует сигнальные каскады реакций, включающих ингибирование фосфатидилинозитид-3 киназы (PI3K) и протеинкиназы В (Akt-киназы) и активацию 3β-киназы гликогенсинтазы (GSK-3β), которая индуцирует гиперфосфорилирование тау-белка, накопление Аβ-олигомеров и окислительный стресс, приводящий к митохондриальной дисфункции, апоптозу клеток, секреции провоспалительных цитокинов и нейродегенерации [7].
Кроме того, нарушение эффектов инсулина может вызвать или усилить процессы нейровоспаления, способствующие развитию нейродегенерации [8]. Нейровоспаление характеризуется активацией микроглии и астроцитов, резко возрастающей продукцией провоспалительных цитокинов (IL-1, IL-6, TNF), формированием внутриклеточных мультибелковых комплексов (инфламмасом), способствующих формированию синаптической и когнитивной дисфункции [9]. Инфламмасомы активируют каспазы 1 и 5, которые запускают секрецию провоспалительных цитокинов (IL-1, IL-18, IL-33), инициируя нейровоспаление c последующим возникновением когнитивных расстройств и слабоумия [10].
Имеются данные о существенном вкладе инфламмасом в развитие инсулинорезистентности, что предполагает существование связи между формированием инфламмасом в клетках головного мозга и локальной инсулинорезистентностью при нейровоспалении и нейродегенерации [11]. Согласно предложенной гипотезе, нарушение церебрального метаболизма глюкозы, связанное с инсулинорезистентностью, запускает каскад патологических реакций, а именно митохондриальную дисфункцию, окислительный стресс, эксайтотоксичность, апоптоз, активацию провоспалительных цитокинов, и, следовательно, способствует накоплению β-амилоида и гиперфосфорилированию тау-белка, тем самым вызывая когнитивную дисфункцию и нейродегенерацию [12].
Одним из потенциальных молекул-маркеров инсулинорезистентности является инсулинрегулируемая аминопептидаза (IRAP). Эта цинк-содержащая металлопептидаза обеспечивает протеолиз нейропептидов (в частности, окситоцина и вазопрессина) [13]. Ее ингибирование улучшает память у экспериментальных животных [14].
Таким образом, БА представляет собой, по-видимому, нейроэндокринное расстройство, связанное с мозгоспецифическим нарушением инсулин/IGF-сигнальных механизмов, т. е. «диабет 3-го типа». Однако молекулярный механизм нарушения передачи сигналов инсулина в нейронах остается в значительной степени неизвестным.
Цель настоящей работы — изучение экспрессии молекул-маркеров, а именно, IRAP, GLUT4 и IL-18 в различных областях головного мозга (гиппокамп, обонятельная луковица) у крыс с экспериментальной БА.
Материал и методы
Моделирование нейродегенерацииКрысам опытной группы (10 самцов в возрасте 7 мес) с помощью стереотаксической установки (Narishige, Япония) в СА1 зону гиппокампа с каждой стороны вводили по 5 мкл бета-амилоида 1−42 (Sigma-Aldrich, США).
Оценку признаков БА начинали с 10-х суток после оперативного вмешательства [18].
Иммуногистохимическое исследованиеНа 14-й день после операции осуществляли транскардиальную перфузию 4% параформальдегидом с последующим забором головного мозга. Мозг фиксировали в 4% нейтральном забуференном формалине, после чего погружали в 20% раствор сахарозы и хранили при +4 °С. С помощью микротома Thermo Scientific Microm HM 650 готовили срезы толщиной 50 мкм. Экспрессию IRAP, GLUT4 и IL-18 изучали методом непрямой иммуногистохимии для свободно плавающих срезов [19]. После промывки в PBS срезы блокировали 3% бычьим сывороточным альбумином (BSA) в PBS и 1% Triton X-100 в течение 1 ч при комнатной температуре с последующим инкубированием в течение ночи с первичными антителами к IRAP (Santa Cruz Biotechnology, sc-365300, rabbit monoclonal) 1:1000, GLUT4 (Abcam, ab654, rabbit monoclonal) 1:10000, IL-18 (Santa Cruz, sc-7954, rabbit polyclonal), NeuN (Millipore, ABN78, rabbit polyclonal) 1:1000 с 3% BSA в PBS и 0,2% Triton X-100 при 4 °C. После инкубации с первичными антителами срезы промывали и инкубировали со вторичными антителами Alexa Conjugated antibody в разведении 1:1000 в течение 2 ч при комнатной температуре.
С помощью микротома Thermo Scientific Microm HM 650 готовили срезы толщиной 50 мкм. Экспрессию IRAP, GLUT4 и IL-18 изучали методом непрямой иммуногистохимии для свободно плавающих срезов [19]. После промывки в PBS срезы блокировали 3% бычьим сывороточным альбумином (BSA) в PBS и 1% Triton X-100 в течение 1 ч при комнатной температуре с последующим инкубированием в течение ночи с первичными антителами к IRAP (Santa Cruz Biotechnology, sc-365300, rabbit monoclonal) 1:1000, GLUT4 (Abcam, ab654, rabbit monoclonal) 1:10000, IL-18 (Santa Cruz, sc-7954, rabbit polyclonal), NeuN (Millipore, ABN78, rabbit polyclonal) 1:1000 с 3% BSA в PBS и 0,2% Triton X-100 при 4 °C. После инкубации с первичными антителами срезы промывали и инкубировали со вторичными антителами Alexa Conjugated antibody в разведении 1:1000 в течение 2 ч при комнатной температуре.
Изображения были получены с помощью конфокального микроскопа Olympus FV 10i. В срезах головного мозга подсчитывали количество клеток нейрональной природы, экспрессирующих маркеры инсулинорезистентности на различных уровнях в гиппокампе (DG) и ольфакторной луковице (OB). Оценивали семь полей зрения для DG и пять полей зрения для OB.
Оценивали семь полей зрения для DG и пять полей зрения для OB.
Статистический анализ результатов проводили с использованием пакета анализа программы MS Excel 2010. В пределах каждой выборки определяли среднее арифметическое и ошибку среднего. Сравнение средних осуществляли с помощью Т-теста при уровне значимости
Результаты
Срезы головного мозга окрашивали по стандартному протоколу двойного непрямого метода иммуногистохимии для оценки экспрессии IRAP, GLUT4 и IL-18 в клетках нейрональной природы (маркер постмитотических нейронов — NeuN). Все измерения проводились в гранулярном слое клеток зубчатой извилины и ольфакторной луковице. При экспериментальной БА отмечалось значимое увеличение экспрессии IL-18 (p=0,001) в гиппокампе по сравнению с контролем (рис.
Таблица 1. Количество клеток (в %), экспрессирующих IL-18 у крыс опытной и контрольной групп
Рис. 1. Экспрессия IL-18 (красный) на нейронах (зеленый) в гиппокампе крыс при экспериментальной болезни Альцгеймера. Колокализация IL-18 с NeuN (желтый).
Несколько иная ситуация наблюдалась в отношении инсулинзависимого глюкозного транспортера GLUT4 (рис. 2, a, б). При Б.А. его экспрессия в гиппокампе (p=0,037) и в ольфакторной луковице (
Таблица 2. Количество клеток (в %), экспрессирующих GLUT4 у крыс опытной и контрольной групп
Рис. 2. (а, б). Экспрессия GLUT4 (красный) на нейронах (зеленый) в гиппокампе крыс при экспериментальной болезни Альцгеймера. Колокализация GLUT4 с NeuN (желтый).
В зубчатой извилине гиппокампа крыс с моделью БА имела место тенденция к увеличению экспрессии IRAP (рис. 3, a, б) в клетках нейрональной природы по сравнению с контролем (p=0,203). Увеличение экспрессии IRAP у подопытных крыс было выявлено также в ольфакторной луковице (табл. 3). Это соответствует нарушению экспрессии GLUT4 при БА [24].
Таблица 3. Количество клеток (в %), экспрессирующих IRAP у крыс опытной и контрольной групп
Рис. 3. (а, б). Экспрессия IRAP (красный) на нейронах (зеленый) в гиппокампе крыс при экспериментальной болезни Альцгеймера. Колокализация IRAP с NeuN (желтый).
Таким образом, в гиппокампе и ольфакторной луковице животных с экспериментальной БА интенсивность экспрессии инсулинрегулируемой аминопептидазы IRAP и провоспалительного цитокина IL-18 возрастает, тогда как экспрессия инсулинзависимого глюкозного транспортера GLUT4 в данных областях головного мозга снижается.
Обсуждение
В мозге больных БА выявлены нейрофибриллярные изменения, потеря нейронов, синаптическая дисфункция как свидетельство эксайтотоксичности, а также выраженный воспалительный процесс. Воспаление играет важную роль в этиопатогенезе БА [25], хотя пока не ясно, является ли оно причиной или вторичным проявлением данного заболевания [26]. Кроме того, остается открытым вопрос о связи воспаления и инсулинорезистентности при нейродегенерации.
Установлено, что резистентность мозга к инсулину возрастает за счет накопления β-амилоида: у пациентов с БА обнаруживается высокая концентрация провоспалительных цитокинов в спинномозговой жидкости [27]. In vivo показано, что воспаление сопровождается образованием и накоплением β-амилоида [21]. β-Амилоид активирует микроглию и астроциты, которые в свою очередь запускают процесс высвобождения провоспалительных цитокинов, ингибирующих рецепторы инсулина за счет увеличения форфорилирования серина IRS-1 (insulin substrate-1) и протеинкиназы В (Akt-киназы), что способствует развитию нейродегенерации [28].
Наши данные [29] о повышении экспрессии провоспалительного цитокина IL-18 на клетках нейрональной природы в гиппокампе и ольфакторной луковице животных с экспериментальной БА могут свидетельствовать о запуске воспалительного процесса и согласуются с результатами исследования экспрессии IL-18 в мозге больных БА.
IL-18 увеличивает экспрессию GSK-3 и циклинзависимой киназы 5 (Cdk5), которые участвуют в гиперфосфорилировании тау-белка [30]. Кроме того, IL-18 может активировать JNK и МАРК р38 сигнальных путей, способствуя запуску апоптоза клеток [31].
Таким образом, IL-18 — один из ключевых медиаторов воспаления и иммунной реакции может играть важную роль в развитии патофизиологических процессов в мозге и способствовать прогрессированию БА.
Доставка глюкозы к клеткам головного мозга осуществляется с помощью глюкозных транспортеров (GLUT). Один из них, инсулинозависимый GLUT4, совместно с рецепторами инсулина в большом количестве экспрессируется в гиппокампе. Инсулин стимулирует экспрессию гена GLUT4 и его транспорт из цитоплазмы к плазматической мембране, тем самым модулируя поглощение и утилизацию глюкозы. Таким образом, инсулин/IGF — сигнальный путь играет ключевую роль в регуляции трансмембранного транспорта глюкозы [12]. Снижение экспрессии GLUT4 в гиппокампе и ольфакторной луковице при БА [32] может быть следствием снижения уровня инсулина в клетках головного мозга [22] и участвовать в нарушении церебрального метаболизма глюкозы.
Инсулин стимулирует экспрессию гена GLUT4 и его транспорт из цитоплазмы к плазматической мембране, тем самым модулируя поглощение и утилизацию глюкозы. Таким образом, инсулин/IGF — сигнальный путь играет ключевую роль в регуляции трансмембранного транспорта глюкозы [12]. Снижение экспрессии GLUT4 в гиппокампе и ольфакторной луковице при БА [32] может быть следствием снижения уровня инсулина в клетках головного мозга [22] и участвовать в нарушении церебрального метаболизма глюкозы.
Выявленная нами тенденция к увеличению экспрессии инсулинрегулируемой аминопептидазы IRAP в клетках нейрональной природы при экспериментальной БА может быть связана с нарушением стимулируемого инсулином транспорта GLUT4 к клеточной мембране, что необходимо для захвата глюкозы клетками мозга [33].
Клинически БА характеризуется прогрессивными нарушениями когнитивных функций и памяти. Показано, что пептиды ангиотензин IV и LVV-геморфин 7, играющие важную роль в процессе обучения и памяти, обладают высоким сродством к IRAP и являются его конкурентными ингибиторами [34]. Высокие концентрации IRAP присутствуют в областях мозга, ответственных за память, а именно в гиппокампе, миндалине и коре [35]. Кроме того, IRAP колоколизован с GLUT4 и регулирует его внутриклеточный транспорт [36].
Высокие концентрации IRAP присутствуют в областях мозга, ответственных за память, а именно в гиппокампе, миндалине и коре [35]. Кроме того, IRAP колоколизован с GLUT4 и регулирует его внутриклеточный транспорт [36].
Известно, что транслокация GLUT4 в мембрану клетки обеспечивается активностью IRAP [37]; отсутствие экспрессии IRAP сопровождается снижением экспрессии и GLUT4 [38]. Поэтому обнаруженное нами несоответствие между экспрессией этих молекул может объясняться отсутствием влияния других инсулинопосредованных [39] или цитоскелет-зависимых [40] механизмов регуляции транспорта GLUT4 в очаге нейровоспаления. В последние годы IRAP считают одним из маркеров инсулинорезистентности при БА. В исследованиях in vivo установлено, что ингибиторы IRAP улучшают память у грызунов [15]. Увеличение экспрессии IRAP и IL-18 в клетках нейрональной природы при нейротоксическом действии β-амилоида соответствует нарушению внутриклеточного транспорта GLUT4 и определяет снижение чувствительности к действию инсулина, что может иметь своим результатом развитие когнитивного дефицита.
Заключение
Уменьшение экспрессии GLUT4 на фоне увеличения экспрессии IRAP и IL-18 в нейронах головного мозга при экспериментальной болезни Альцгеймера свидетельствует о сопряженности механизмов нейровоспаления и инсулинорезистентности, что открывает возможность новых подходов к направленной фармакологической коррекции нейродегенерации.
Информация о финансировании и конфликте интересовУчастие авторов:
Концепция и дизайн исследования —Комлева Ю.К..
Сбор и обработка материала — Горина Я.В., Комлева Ю.К., Лопатина О.Л., Волкова В.В.
Статистическая обработка данных — Герцог Г.Е., Попова Н.Н.
Написание текста — Горина Я.В..
Редактирование — Салмина А.Б..
Работа выполнена при поддержке гранта Президента Р.Ф. для ведущих научных школ РФ (НШ-1172.2014.7).
Авторы декларируют отсутствие явных и потенциальных конфликтов интересов.
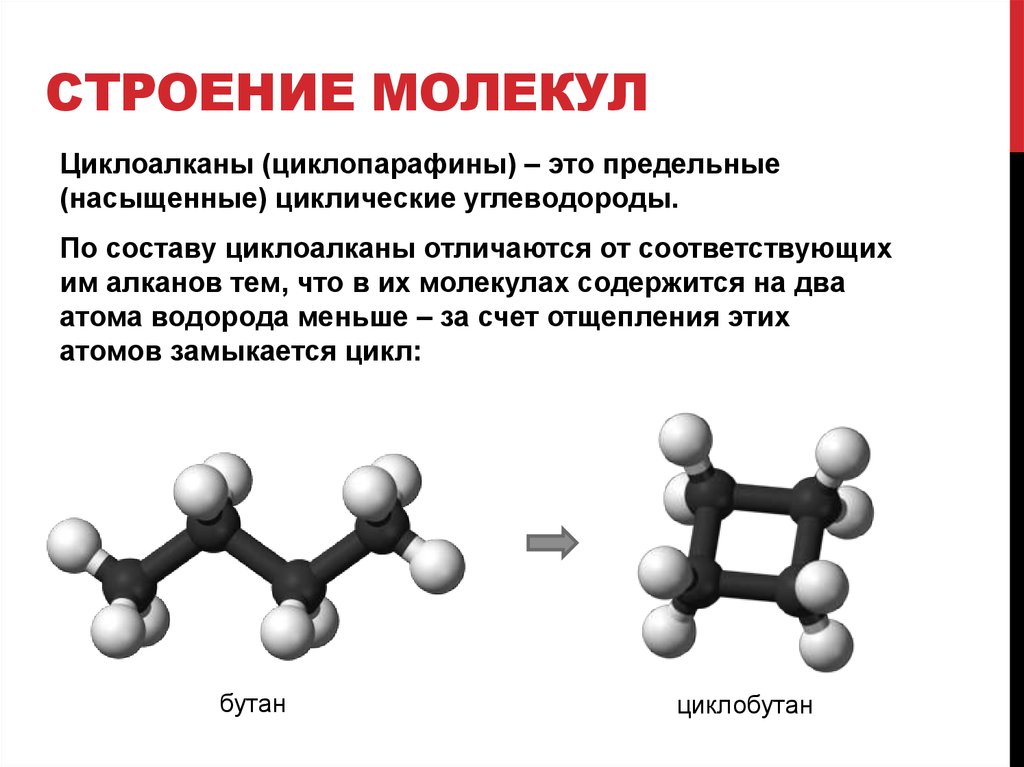
Полимеризация — Что такое Полимеризация?
62548
Полимеризация — процесс образования высокомолекулярного вещества путем многократного присоединения молекул
Полимеризация — процесс образования высокомолекулярного вещества (полимера) путем многократного присоединения молекул низкомолекулярного вещества (мономера, олигомера) к активным центрам в растущей молекуле полимера.
Молекула мономера, входящая в состав полимера, образует так называемое мономерное (структурное) звено.
Элементный состав (молекулярные формулы) мономера и полимера приблизительно одинаков.
Мономерами являются соединения, содержащие кратные связи, которые способны, раскрываясь, образовывать новые связи с другими молекулами, обеспечивая рост цепей.
Механизм полимеризации обычно включает в себя ряд связанных стадий:
- инициирование — зарождение активных центров полимеризации;
- рост (продолжение) цепи — процесс последовательного присоединения молекул мономеров к центрам;
- передача цепи — переход активного центра на другую молекулу;
- разветвление цепи — образование нескольких активных центров из одного;
- обрыв цепи — гибель активных центров.
Виды полимеризации
В основу классификации полимеризации могут быть положены различные признаки:
число типов молекул мономеров:
- гомополимеризация — полимеризация одинаковых мономеров;
- сополимеризация — полимеризация двух и более разных мономеров.
- природа активного центра и механизм процесса:
- радикальная полимеризация — активными центрами являются свободные радикалы;
-
ионная полимеризация — активные центры ионы или поляризованные молекулы.


Фазовое состояние мономеров:
- газофазная полимеризация;
- жидкофазная полимеризация;
- твердофазная полимеризация.
Структура области, в которой сосредоточены активные центры:
- объемная полимеризация — полимеризация во всем объеме мономера;
- фронтальная полимеризация — экзотермическая полимеризация в узком фронте, распространяющемся в среде мономера;
- эмульсионная полимеризация — полимеризация на поверхности высокодиспергированных частиц мономера в эмульсии.
Способ инициирования:
- фотополимеризация;
- термическая полимеризация;
- радиационная полимеризация и др.
Структурные особенности полученного полимера:
- стереорегулярная полимеризация — полимеризация с образованием полимеров с упорядоченной пространственной структурой.

Технологические особенности полимеризации:
- полимеризация при высоком давлении и др.
Химическая природа мономеров:
В основе химических превращений полимеров лежит замена одних функциональных групп на другие, что проходит без изменения степени полимеризации.
Последние новости
Рисунки и данные в Идентификация молекулярных особенностей, которые связаны с биологической функцией внутренне неупорядоченных белковых областей
5 рисунков и 2 дополнительных файла
Рисунки
Схема статистической модели FAIDR.
Примеры входных данных (функция, молекулярные признаки) и выходных данных (вероятность FAIDR, связь с функцией) показаны. Учитывая набор признаков (например, молекулярные признаки, включающие эволюционные сигнатуры, как показано) для одного или нескольких IDR в данном белке и информацию о функциональной аннотации (например, аннотирован ли белок функцией связывания мРНК или функцией ядрышка, как показано ), FAIDR выводит вероятность того, что IDR связан с данной функцией, а также силу и направление связи с функцией для каждого молекулярного признака. Молекулярные признаки, включающие в себя эволюционные сигнатуры, представлены с помощью масштабирования тепловой карты от синего (уменьшение среднего значения или дисперсии признака по сравнению с нашим нулевым ожиданием эволюции IDR) до желтого (увеличение среднего значения или дисперсии признака по сравнению с нашим нулевым ожиданием развития IDR). . Связь с функцией представлена тепловой картой, масштабируемой от синего (отрицательная ассоциация) до желтого (положительная связь). Например, для функции ядрышка (как показано) средние консенсусные сайты CKII сильно положительно связаны с этой функцией (обозначены желтым цветом), тогда как для связывания мРНК (также показано) среднее содержание треонина отрицательно связано с этой функцией (обозначено синим цветом). ).
Молекулярные признаки, включающие в себя эволюционные сигнатуры, представлены с помощью масштабирования тепловой карты от синего (уменьшение среднего значения или дисперсии признака по сравнению с нашим нулевым ожиданием эволюции IDR) до желтого (увеличение среднего значения или дисперсии признака по сравнению с нашим нулевым ожиданием развития IDR). . Связь с функцией представлена тепловой картой, масштабируемой от синего (отрицательная ассоциация) до желтого (положительная связь). Например, для функции ядрышка (как показано) средние консенсусные сайты CKII сильно положительно связаны с этой функцией (обозначены желтым цветом), тогда как для связывания мРНК (также показано) среднее содержание треонина отрицательно связано с этой функцией (обозначено синим цветом). ).
Вероятностная модель FAIDR.
Незакрашенные кружки представляют скрытые переменные, а серые кружки – известные переменные. Функции или фенотипы рассматриваются как бинарные переменные, где Y = 1, если белок связан с этой функцией или фенотипом, и Y = 0, если белок не связан. Для i й белок, у нас есть набор признаков, Z i для каждой из r i неупорядоченных областей. Мы используем скрытую переменную X i для представления IDR, отвечающего за функцию, так что если X ij = 1, j -й IDR в i -м белке равен тот, кто отвечает за функцию. Повсюду мы использовали экспоненциальный априор, λ = 0,2 для коэффициентов, b (эквивалентно постоянному штрафу L1), который, как мы обнаружили, ограничивает переобучение для наших функций с небольшим количеством положительных результатов.
Функции или фенотипы рассматриваются как бинарные переменные, где Y = 1, если белок связан с этой функцией или фенотипом, и Y = 0, если белок не связан. Для i й белок, у нас есть набор признаков, Z i для каждой из r i неупорядоченных областей. Мы используем скрытую переменную X i для представления IDR, отвечающего за функцию, так что если X ij = 1, j -й IDR в i -м белке равен тот, кто отвечает за функцию. Повсюду мы использовали экспоненциальный априор, λ = 0,2 для коэффициентов, b (эквивалентно постоянному штрафу L1), который, как мы обнаружили, ограничивает переобучение для наших функций с небольшим количеством положительных результатов.
FAIDR, обученный на эволюционных сигнатурах IDR, может предсказывать невидимые данные по сравнению с современными, специфическими предикторами сигналов митохондриального нацеливания (Mitofates [Fukasawa et al.
 , 2015]) и субстратов Cdc28 (Condens [Lai et al., 2012]).
, 2015]) и субстратов Cdc28 (Condens [Lai et al., 2012]).Рабочие кривые приемника (ROC) на протянутой 20% выборке показаны для FAIDR, обученного на эволюционных сигнатурах (синие), по сравнению с Mitofates ( A ) и Condens ( B ) (оранжевый). Площадь под кривой (AUC) указана в легенде справа внизу для каждого метода. Количество положительных РДЭ (Np) в оставшихся 20% указано в левом верхнем углу каждого графика.
Рисунок 2—исходные данные 1
Данные для кривых ROC, включая координаты IDR, вероятности FAIDR для вытянутых IDR и наземные метки истинности из экспериментальных данных, сопоставленных с каждым IDR.
idr_name_protein_coordinates.txt определяет имена IDR как координаты аминокислот в каждом белке. holdout_idr_probability_cdc28_mito.csv — это вероятность FAIDR для РДЭ, которые не были включены в обучающий набор («отложенные РДЭ»).
total_idr_expt_db_overlap_lai_2009_cdc28.csv представляет собой двоичную сводку того, фосфорилируется ли каждый IDR с помощью Cdc28 в соответствии с экспериментальными данными, сопоставленными из координат белка с каждым IDR (данные, собранные Lai et al., 2012). total_idr_expt_db_overlap_vogtle_2009_mitochondrial_nterm.csv представляет собой бинарную сводку того, является ли каждый IDR митохондриальным N-концевым нацеливающим сигналом в соответствии с экспериментальными данными, сопоставленными с белковыми координатами для каждого IDR (на основе экспериментального исследования всего протеома, проведенного Vögtle et al., 2009).- https://cdn.elifesciences.org/articles/60220/elife-60220-fig2-data1-v2.zip
Скачать elife-60220-fig2-data1-v2.zip
 total_idr_expt_db_overlap_lai_2009_cdc28.csv представляет собой двоичную сводку того, фосфорилируется ли каждый IDR с помощью Cdc28 в соответствии с экспериментальными данными, сопоставленными из координат белка с каждым IDR (данные, собранные Lai et al., 2012). total_idr_expt_db_overlap_vogtle_2009_mitochondrial_nterm.csv представляет собой бинарную сводку того, является ли каждый IDR митохондриальным N-концевым нацеливающим сигналом в соответствии с экспериментальными данными, сопоставленными с белковыми координатами для каждого IDR (на основе экспериментального исследования всего протеома, проведенного Vögtle et al., 2009).
total_idr_expt_db_overlap_lai_2009_cdc28.csv представляет собой двоичную сводку того, фосфорилируется ли каждый IDR с помощью Cdc28 в соответствии с экспериментальными данными, сопоставленными из координат белка с каждым IDR (данные, собранные Lai et al., 2012). total_idr_expt_db_overlap_vogtle_2009_mitochondrial_nterm.csv представляет собой бинарную сводку того, является ли каждый IDR митохондриальным N-концевым нацеливающим сигналом в соответствии с экспериментальными данными, сопоставленными с белковыми координатами для каждого IDR (на основе экспериментального исследования всего протеома, проведенного Vögtle et al., 2009).Прогнозирование различных функций и фенотипов белков с помощью FAIDR.

Показана средняя площадь под рабочей кривой приемника (AUC), рассчитанная с использованием ROCR (Sing et al., 2009) на отложенных 20 % при пятикратной перекрестной проверке, рассчитанная с использованием 82 молекулярных признаков на Saccharomyces cerevisiae. 9Только последовательность IDR 0018 (незакрашенные столбцы) или по эволюционным сигнатурам (закрашенные столбцы, см. Текст). Столбики погрешностей представляют собой стандартное отклонение. Функциональные аннотации взяты из GO (получены из SGD [Cherry et al., 2012]), за исключением фенотипов делеции генов (нежизнеспособный, респираторный рост, накопление РНК, из SGD), субстратов Cdc28 (yeastKID, Sharifpoor et al., 2011), мРНП (Mitchell et al., 2013) и обратимая тепловая агрегация (Wallace et al., 2015).
Молекулярные характеристики IDR связаны со специфическими функциями.
( A ) Иерархическая кластеризация t-статистики, полученная в результате регрессии 23 функций и фенотипов по эволюционным признакам (средние значения и дисперсии 82 молекулярных признаков). Символы рядом с именами функций/фенотипов указывают на связанные функции/фенотипы. Указанные подгруппы ( i–ix ) упоминаются и описываются в основном тексте. ( Б ) Примеры положительных и отрицательных прогностических молекулярных признаков для передачи сигналов ( i ) и PMLO (vi-ix).
Рисунок 3—исходные данные 1
Необработанная T-статистика, используемая для кластерного анализа и отображения.
Оставшиеся молекулярные признаки (n = 73) и связанные функции/фенотипы (n = 23) затем были иерархически сгруппированы с использованием взвешенного нецентрированного корреляционного расстояния («кластер» и «вычисление весов» отмечены флажками с отсечкой = 0,1 и показателем степени = 1).
на кластере 3.0, используя среднюю связь. Кластерную t-статистику, лежащую в основе рисунка 3 (tstat_table_atleast1_over_3_v2.cdt), можно просмотреть/исследовать с помощью Java Treeview.- https://cdn.elifesciences.org/articles/60220/elife-60220-fig3-data1-v2.zip
Скачать elife-60220-fig3-data1-v2.zip
 на кластере 3.0, используя среднюю связь. Кластерную t-статистику, лежащую в основе рисунка 3 (tstat_table_atleast1_over_3_v2.cdt), можно просмотреть/исследовать с помощью Java Treeview.
на кластере 3.0, используя среднюю связь. Кластерную t-статистику, лежащую в основе рисунка 3 (tstat_table_atleast1_over_3_v2.cdt), можно просмотреть/исследовать с помощью Java Treeview.Молекулярные особенности, которые, по прогнозам, связаны с аннотацией GO «митохондрия», влияют на фенотип митохондриальной локализации.
( A ) Вектор признаков Z-показателей (эволюционная сигнатура) N-концевого IDR Cox15 (Cox15 IDR 1) (ось y) нанесен на график относительно прогностических признаков (t-статистика) для митохондриальной функции, как определено FAIDR (ось x). Двумя главными признаками, связанными с митохондриальной локализацией, являются средняя изоэлектрическая точка (pI), обведенная красным, и средняя гидрофобность (H), обведенная синим. ( B ) Аминокислотные последовательности для каждого варианта IDR Cox15. Последовательности IDR дикого типа и смоделированные Cox15 IDR сравнивают с той же последовательностью с мутациями, изменяющими изоэлектрическую точку (pI) и гидрофобность. Последовательности с вариабельными остатками (серые) визуализируются с помощью Jalview (Waterhouse et al., 2009).). Также показаны прогнозы Mitofates (Fukasawa et al., 2015), предсказателя последовательностей, нацеленных на митохондрии: прогноз того, является ли данная последовательность митохондриальной предварительной последовательностью (галочка), а также предсказанное местоположение сайт расщепления в последовательностях дикого типа и мутантных последовательностях (красный прямоугольник), области, для которых прогнозируется высокий (фиолетовый прямоугольник) или низкий (розовый прямоугольник) показатель максимальной положительно заряженной амфифильности, и область мотива распознавания TOM20 (рецептор) (зеленый коробка).
Двумя главными признаками, связанными с митохондриальной локализацией, являются средняя изоэлектрическая точка (pI), обведенная красным, и средняя гидрофобность (H), обведенная синим. ( B ) Аминокислотные последовательности для каждого варианта IDR Cox15. Последовательности IDR дикого типа и смоделированные Cox15 IDR сравнивают с той же последовательностью с мутациями, изменяющими изоэлектрическую точку (pI) и гидрофобность. Последовательности с вариабельными остатками (серые) визуализируются с помощью Jalview (Waterhouse et al., 2009).). Также показаны прогнозы Mitofates (Fukasawa et al., 2015), предсказателя последовательностей, нацеленных на митохондрии: прогноз того, является ли данная последовательность митохондриальной предварительной последовательностью (галочка), а также предсказанное местоположение сайт расщепления в последовательностях дикого типа и мутантных последовательностях (красный прямоугольник), области, для которых прогнозируется высокий (фиолетовый прямоугольник) или низкий (розовый прямоугольник) показатель максимальной положительно заряженной амфифильности, и область мотива распознавания TOM20 (рецептор) (зеленый коробка). ( C ) Микрофотографии, показывающие фенотип митохондриальной локализации для различных штаммов почкующихся дрожжей, которые различаются по своим N-концевым IDR Цокс15. Зеленым цветом показана локализация Cox15, помеченная GFP. Слева направо: IDR дикого типа, IDR Cox15 с низким pI, IDR Cox15 с низкой гидрофобностью, смоделированный IDR, смоделированный IDR с высоким pI. Масштабная линейка представляет 1 мкм.
( C ) Микрофотографии, показывающие фенотип митохондриальной локализации для различных штаммов почкующихся дрожжей, которые различаются по своим N-концевым IDR Цокс15. Зеленым цветом показана локализация Cox15, помеченная GFP. Слева направо: IDR дикого типа, IDR Cox15 с низким pI, IDR Cox15 с низкой гидрофобностью, смоделированный IDR, смоделированный IDR с высоким pI. Масштабная линейка представляет 1 мкм.
Микрофотографии, показывающие (столбцы слева направо) наложение в светлом поле, DAPI, Cox15-GFP и Cox15-GFP/DAPI с 4-кратным увеличением различных штаммов почкующихся дрожжей с различными генотипами Cox15 IDR (метки в левой части изображения).
DAPI окрашивает ДНК в ядре и митохондриях. Изображения штаммов Cox15 WT и Cox15 sim IDR с высоким pI показывают, что Cox15-GFP локализован в митохондриях в этих штаммах, в то время как Cox15-GFP не локализован в митохондриях в Cox15 sim IDR, Cox15 IDR с низким pI и Cox15 IDR с низкой гидрофобностью. Белые прямоугольники обозначают увеличенные в 4 раза области, показанные в крайнем правом столбце. Белые стрелки указывают области, где происходит перекрытие неядерных DAPI и Cox15-GFP (т.е. митохондриальная локализация).
Белые прямоугольники обозначают увеличенные в 4 раза области, показанные в крайнем правом столбце. Белые стрелки указывают области, где происходит перекрытие неядерных DAPI и Cox15-GFP (т.е. митохондриальная локализация).
Идентификация специфической последовательности IDR, связанной с активностью фактора транскрипции в неохарактеризованном белке.
( A ) Гистограмма вероятностей ассоциации IDR с функцией связывания ДНК, специфичной для последовательности (фактор транскрипции или «TF»). Указаны лучшие прогнозы. Среди основных прогнозов IDR известных факторов транскрипции отмечены зеленой точкой. Mfg1 представляет собой неохарактеризованный белок. ( B ) Белок Mfg1 координируется со всеми известными аннотациями доменов (посредством PFAM), прогнозированием нарушений (посредством DISOPRED3) и вероятностью FAIDR для ассоциации с указанным связыванием ДНК, специфичным для последовательности. ( C ) Вектор признаков (эволюционная сигнатура) для Mfg1 IDR 1 (ось ординат) нанесен в зависимости от прогностических признаков для функции связывания ДНК, специфичной для фактора транскрипции/последовательности, как определено с помощью FAIDR (ось абсцисс). Логарифмическая дисперсия остатков глутамина (Q) выделена как одна из основных характеристик (оранжевый кружок), связанных с функцией TF. Остатки глутамина (Q) указаны (оранжевым цветом) в IDR Mfg1 и ортологов, полученных из YGOB (Byrne and Wolfe, 2005). Все остальные (не глютаминовые) остатки обозначены черным цветом. Множественное выравнивание последовательностей ортологов визуализируется с помощью Jalview (Waterhouse et al., 2009).). Species in alignment (from bottom to top) are as follows: Saccharomyces cerevisiae , Saccharomyces mikatae , Saccharomyces kudriavzevii , Saccharomyces uvarum , Candida glabrata , Kazachstania africana , Kazachstania naganishii , Naumovozyma castellii , Naumovozyma dairenensis , Zygosaccharomyces rouxii , Torulaspora delbrueckii , Eremothecium (Ashbya) gossypii , Eremothecium (Ashbya) cymbalariae , Lachancea kluyveri , Lachancea thermotolerans и Lachancea waltii .
( C ) Вектор признаков (эволюционная сигнатура) для Mfg1 IDR 1 (ось ординат) нанесен в зависимости от прогностических признаков для функции связывания ДНК, специфичной для фактора транскрипции/последовательности, как определено с помощью FAIDR (ось абсцисс). Логарифмическая дисперсия остатков глутамина (Q) выделена как одна из основных характеристик (оранжевый кружок), связанных с функцией TF. Остатки глутамина (Q) указаны (оранжевым цветом) в IDR Mfg1 и ортологов, полученных из YGOB (Byrne and Wolfe, 2005). Все остальные (не глютаминовые) остатки обозначены черным цветом. Множественное выравнивание последовательностей ортологов визуализируется с помощью Jalview (Waterhouse et al., 2009).). Species in alignment (from bottom to top) are as follows: Saccharomyces cerevisiae , Saccharomyces mikatae , Saccharomyces kudriavzevii , Saccharomyces uvarum , Candida glabrata , Kazachstania africana , Kazachstania naganishii , Naumovozyma castellii , Naumovozyma dairenensis , Zygosaccharomyces rouxii , Torulaspora delbrueckii , Eremothecium (Ashbya) gossypii , Eremothecium (Ashbya) cymbalariae , Lachancea kluyveri , Lachancea thermotolerans и Lachancea waltii .
Пример модульной функции IDR, предсказанной FAIDR.
( A ) Вероятности FAIDR для РДЭ в Ebp2. ( В ) и ( C ) Векторы признаков (эволюционные сигнатуры) для Ebp2 IDR (ось Y), нанесенные на график относительно прогностических признаков для ядрышка ( B ) и связывания мРНК ( C ) (ось x), как определено FAIDR. Основные прогностические признаки ядрышка ( B ) и связывания мРНК ( C ) выделены красным цветом. ( D ) и ( E ) Выравнивание IDR от Ebp2, где сайты консенсусного фосфорилирования CKII ([ST].[DE][DE]) и (отсутствие) треонина (T) выделены черным цветом соответственно. . Стрелки над последовательностью указывают на фосфорилированные остатки (из SGD [Cherry et al., 2012]) в Saccharomyces cerevisiae (верхняя последовательность белка в каждом случае). Оттенок синего представляет собой процент идентичности в множественном выравнивании последовательностей (темно-синий цвет соответствует более высокому проценту идентичности). Ebp2 аннотирован в GO (The Gene Ontology Consortium, 2019) как локализованный в ядрышке и связывающий мРНК. ( D ) и ( E ) Первый IDR показывает обилие консенсусных сайтов фосфорилирования CKII, тогда как второй IDR показывает снижение треонина (T). Примечательно, что эти предсказания согласуются с сообщением о том, что C-концевой укороченный мутант Ebp2, включающий второй IDR (Ionescu et al., 2004), приводит к дефектам, отличным от функций, связанных с ядрышковой рРНК. Этот пример иллюстрирует, как разные IDR в белках с несколькими IDR могут быть связаны с разными функциями: IDR содержат разные молекулярные особенности.
Оттенок синего представляет собой процент идентичности в множественном выравнивании последовательностей (темно-синий цвет соответствует более высокому проценту идентичности). Ebp2 аннотирован в GO (The Gene Ontology Consortium, 2019) как локализованный в ядрышке и связывающий мРНК. ( D ) и ( E ) Первый IDR показывает обилие консенсусных сайтов фосфорилирования CKII, тогда как второй IDR показывает снижение треонина (T). Примечательно, что эти предсказания согласуются с сообщением о том, что C-концевой укороченный мутант Ebp2, включающий второй IDR (Ionescu et al., 2004), приводит к дефектам, отличным от функций, связанных с ядрышковой рРНК. Этот пример иллюстрирует, как разные IDR в белках с несколькими IDR могут быть связаны с разными функциями: IDR содержат разные молекулярные особенности.
Дополнительные файлы
Ссылки для скачивания
Список ссылок, состоящий из двух частей, для загрузки статьи или частей статьи в различных форматах.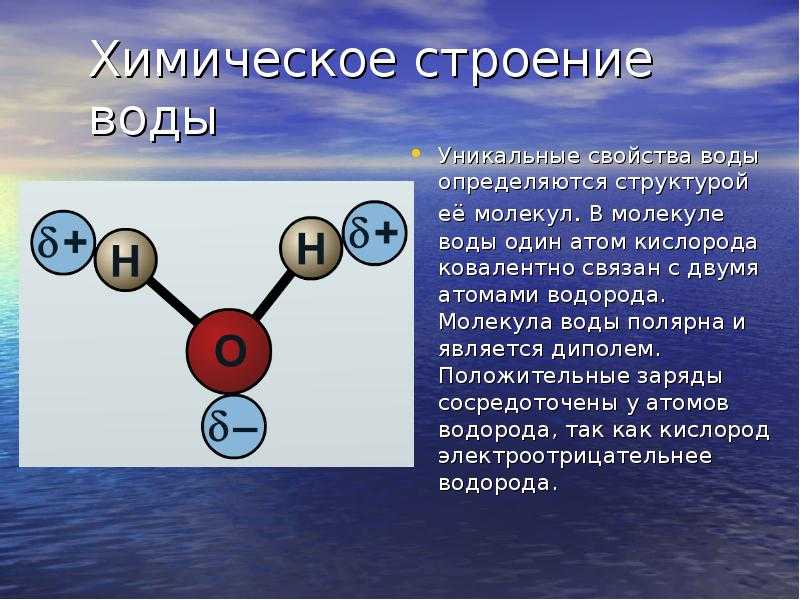
Открытые цитаты (ссылки для открытия цитат из этой статьи в различных онлайн-сервисах управления ссылками)
- Менделей
- ЧитатьКуб»>
Процитировать эту статью (ссылки для загрузки цитат из этой статьи в форматах, совместимых с различными инструментами управления ссылками)
- Таране Зарин
- Боб Стром
- Ган Пэн
- Ива Притишанац
- Джули Д Форман-Кей
- Алан М Моисей
(2021)
Идентификация молекулярных особенностей, связанных с биологической функцией внутренне неупорядоченных белковых участков
eLife 10 :e60220.
https://doi.org/10.7554/eLife.60220
- Скачать БибТекс
- Скачать . RIS
 RIS
RISМолекулярные особенности гидратных слоев, изученные методом атомно-силовой микроскопии
Чжэнцин Чжан, и Сеол Рю, б Юнхо Ан * с и Джункён Джанг * и
Принадлежности автора
* Соответствующие авторы
и Кафедра наноэнергетики, Пусанский национальный университет, Пусан 46241, Южная Корея
Электронная почта: jkjang@pusan. ac.kr
ac.kr
б Кафедра химии, Университет Чосон, Кванджу 61452, Республика Корея
с Школа гуманитарных наук, Корейский технологический и педагогический университет, Чхонан 31253, Республика Корея
Электронная почта: [email protected]
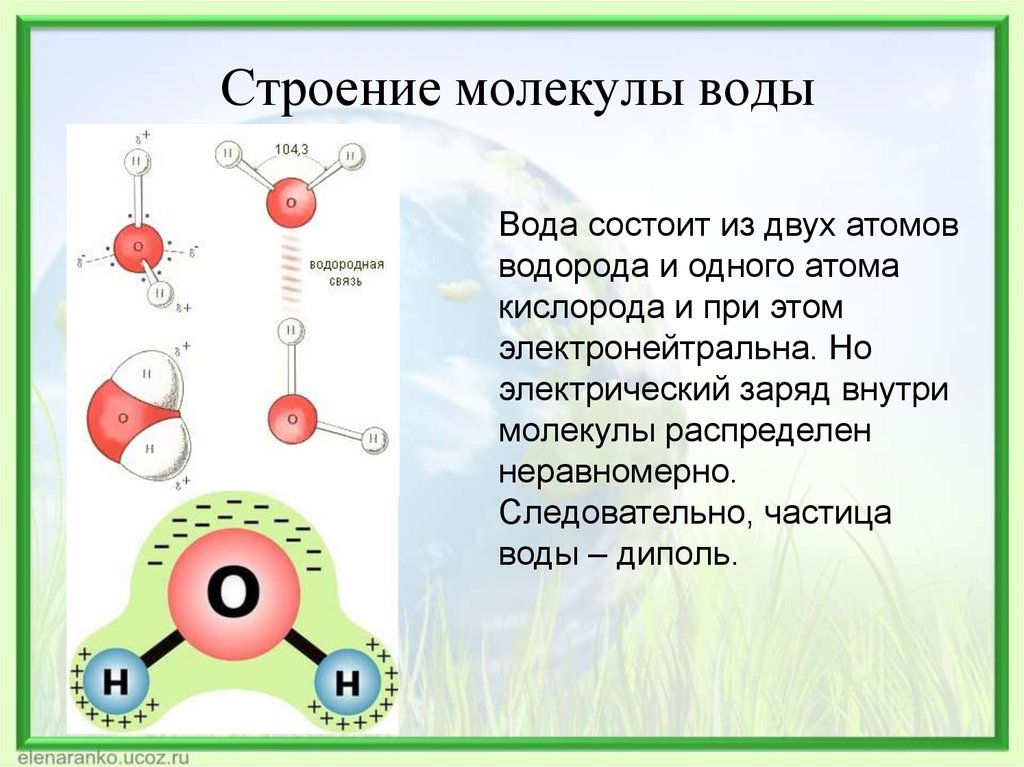
Аннотация
Структурно-упорядоченные слои воды повсеместно образуются на твердой поверхности в водном растворе или в условиях окружающей среды. Хотя такие слои гидратации обычно исследуются с помощью атомно-силовой микроскопии (АСМ) , текущее понимание того, как слои гидратации проявляют себя в эксперименте с АСМ, далеко не полное. Используя моделирование молекулярной динамики, мы исследуем слои гидратации на гидрофильной или гидрофобной поверхности, которые исследуются наноразмерной иглой. Мы изучаем плотность и молекулярную ориентацию воды, свободную энергию и силу, действующую на иглу, изменяя расстояние между иглой и поверхностью. Кривая сила-расстояние колеблется из-за перехода между моно-, би- и трехслойной водой, заключенной между наконечником и поверхностью. Если и наконечник, и поверхность гидрофобны, вода, находящаяся между наконечником и поверхностью, испаряется из-за перехода смачивания, создавая гидрофобную силу без колебаний. Периодичность колебаний силы отличается от структурной периодичности воды. В непосредственной близости от наконечника молекулярные диполи выравниваются параллельно поверхности, независимо от того, являются ли кончик и поверхность гидрофильными или гидрофобными.
Хотя такие слои гидратации обычно исследуются с помощью атомно-силовой микроскопии (АСМ) , текущее понимание того, как слои гидратации проявляют себя в эксперименте с АСМ, далеко не полное. Используя моделирование молекулярной динамики, мы исследуем слои гидратации на гидрофильной или гидрофобной поверхности, которые исследуются наноразмерной иглой. Мы изучаем плотность и молекулярную ориентацию воды, свободную энергию и силу, действующую на иглу, изменяя расстояние между иглой и поверхностью. Кривая сила-расстояние колеблется из-за перехода между моно-, би- и трехслойной водой, заключенной между наконечником и поверхностью. Если и наконечник, и поверхность гидрофобны, вода, находящаяся между наконечником и поверхностью, испаряется из-за перехода смачивания, создавая гидрофобную силу без колебаний. Периодичность колебаний силы отличается от структурной периодичности воды. В непосредственной близости от наконечника молекулярные диполи выравниваются параллельно поверхности, независимо от того, являются ли кончик и поверхность гидрофильными или гидрофобными.