
Виды электрических сигналов
Цель рассказа показать в чем суть понятия «сигнал», какие распространённые сигналы существуют и какие у них общие характеристики.
Что такое сигнал? На этот вопрос даже маленький ребёнок скажет, что это «такая штука, с помощью которой можно что-нибудь сообщить». Например, с помощью зеркала и солнца можно передавать сигналы на расстояние прямой видимости. На кораблях, сигналы когда-то передавали с помощью флажков-семафоров. Занимались этим специально обученые сигнальщики. Таким образом с помощью таких флажков передавалась информация. Вот как можно передать слово «сигнал»:
В природе существует огромное множество сигналов. Да по сути что угодно может быть сигналом: оставленная на столе записка, какой-нибудь звук — могут служить сигналом к началу определённого действия.
Ладно, с такими сигналами всё понятно поэтому перейду к электрическим сигналам, которых в природе не меньше чем любых других. Но их хотя бы можно как-то условно разбить на группы: треугольный, синусоидальный, прямоугольный, пилообразный, одиночный импульс и т. д. Все эти сигналы названы так за то, как они выглядят, если их изобразить их на графике.
д. Все эти сигналы названы так за то, как они выглядят, если их изобразить их на графике.
Сигналы могут быть использованы как метроном для отсчета тактов (в качестве тактирующего сигнала), для отсчета времени, в качестве управляющих импульсов, для управления двигателями или для тестирования оборудования и передачи информации.
Характеристики эл. сигналов
В некотором смысле электрический сигнал — это график, отражающий изменение напряжения или тока с течением времени. Что по-русски означает: если взять карандаш и по оси Х отметить время, а по Y напряжение или ток, и отметить точками соответствующие значения напряжения в конкретные моменты времени, то итоговое изображение будет показывать форму сигнала:
Электрических сигналов очень много, но их можно разбить на две большие группы:
- Однонаправленные
- Двунаправленные
Т.е. в однонаправленных ток течет в одну сторону (либо не течет вообще), а в двунаправленных ток является переменным и протекает то «туда», то «сюда».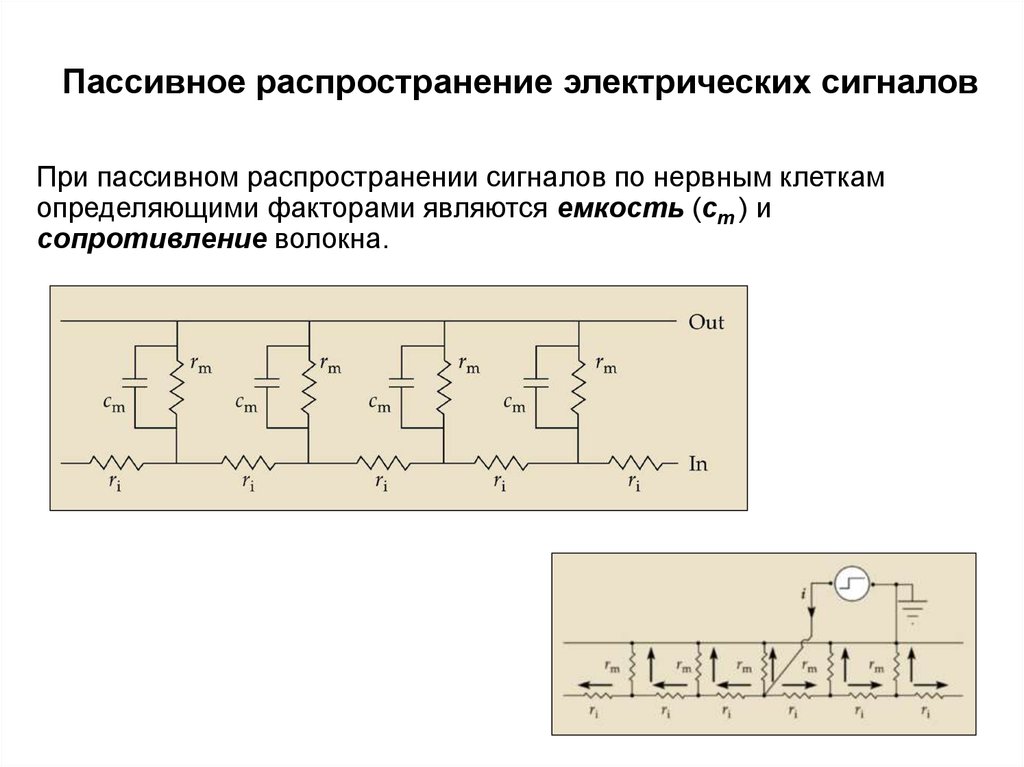
Все сигналы, независимо от типа, обладают следующими характеристиками:
- Период — промежуток времени, через который сигнал начинает повторять себя. Обозначается чаще всего T
- Частота — обозначает сколько раз сигнал повториться за 1 секунду. Измеряется в герцах. К примеру 1Гц = 1 повторение в секунду. Частота является обратным значением периода ( ƒ = 1/T )
- Амплитуда — измеряется в вольтах или амперах (в зависимости от того какой сигнал: ток или напряжение). Амплитуда обозначает «силу» сигнала. Как сильно отклоняется график сигнала от оси Х.
Виды сигналов
СинусоидаДумаю, что представлять функцию, чей график на картинке выше нет смысла — это хорошо тебе известная sin(x). Её период равен 360o или 2pi радиан (2pi радиан =360o).
А если разделить поделить 1 сек на период T, то ты узнаешь сколько периодов укалдывается в 1 сек или, другими словами, как часто период повторяется.
Частота и период обратны друг другу. Чем длинней период, тем меньше частота и наоборот. Связь между частотой и периодом выражается простыми соотношениями:
| Суфикс | Полное значение | Сокращение | Обозначает время |
| Кило | Тысяча (Килогерц) | 1 миллисекунда (10-3) | |
| Мега | Миллион (Мегагерц) | МГц | 1 микросекунда (10-6) |
| Гига | Миллиард (Гигагерц) | ГГц | 1 наносекунда (10-9) |
| Тера | Триллион (Терагерц) | ТГц | 1 пикосекунда (10-12) |
Сигналы, которые по форме напоминают прямоугольники, так и называют «прямоугольные сигналы». Их условно можно разделить на просто прямоугольне сигналы и меандры. Меандр — это прямоугольный сигнал, у которого длительность импульса и паузы равны. А если сложить длительность паузы и импульса, то получим период меандра.
Их условно можно разделить на просто прямоугольне сигналы и меандры. Меандр — это прямоугольный сигнал, у которого длительность импульса и паузы равны. А если сложить длительность паузы и импульса, то получим период меандра.
Обычный прямоугольный сигнал отличается от меандра тем, что имеет разную длительность импульса и паузы (отсутствие импульса). Смотри картинку ниже — она скажет лучше тысячи слов.
Кстати, для прямоугольных сигналов существует еще два термина, которые следует знать. Они обратны друг другу (как период и частота). Это скажность и коээффициент заполнения. Скажность (S)равняется отношению периода к длительности импульса и наоборот для коэфф. заполнения.
Таким образом меандр — это прямоугольный сигнал со скважностью равной 2. Так как у него период в два раза больше длительности импульса.
S — скважность, D — коэффициент заполнения, T — период импульсов, — длительность импульса.
Кстати, на графиках выше показаны идеальные прямоугольные сигналы. В жизни они выглядят слегка иначе, так как ни в одном устройстве сигнал не может измениться абсолютно мгновенно от 0 до какого-то значения и обратно спуститься до нуля.
Треугольный сигналЕсли подняться на гору, а затем сразу спуститься и записать изменение высоты нашего положения на графике, то получим треугольный сигнал. Груое сравнение, но правдивое. В треугольный сигналах напряжение (ток) сначала возрастает, а затем тут же начинает уменьшаться. И для классического треугольного сигнала время возрастания равно времени убывания (и равно половине периода).
Если же у такого сигнала время возрастания меньше или больше времени убывания, то такие сигналы уже называют пилообразными. И о них ниже.
Пилообразный сигнал
Как я уже писал выше, несимметричный треугольный сигнал называется пилообразным. Все эти названи условны и нужны просто для удобства.
Вот такое простое введение в электрические сигналы. В природе их существует множество, но выше описаны те, что в нашем радиолюбительском деле встречаются довольно часто. Надеюсь, что теперь ты будешь больше знать про них.
В природе их существует множество, но выше описаны те, что в нашем радиолюбительском деле встречаются довольно часто. Надеюсь, что теперь ты будешь больше знать про них.
Что почитать дальше
- Юкио Сато. Обработка сигналов
- Каганов. Радиотехические цепи и сгналы
- Как пользоваться осциллографом и для чего он вообще нужен. Часть I
- Как пользоваться осциллографом и для чего он вообще нужен. Часть II
p.s. Часть материала основана на интересной, но многословной статье отсюда. Переводом этот текст не назвать, так что будем считать его вольным пересказом. Картинки адаптированы под русский язык =)
/blog/vidyi-elektricheskih-signalov/
Электрические сигналы бегут по электрическим цепям в радиоприёмниках, усилителях мощности, устройствах на микроконтроллерах и любых других электронных устройствах. Я расскажу какие виды сигналов существуют и какими характеристиками они обладают.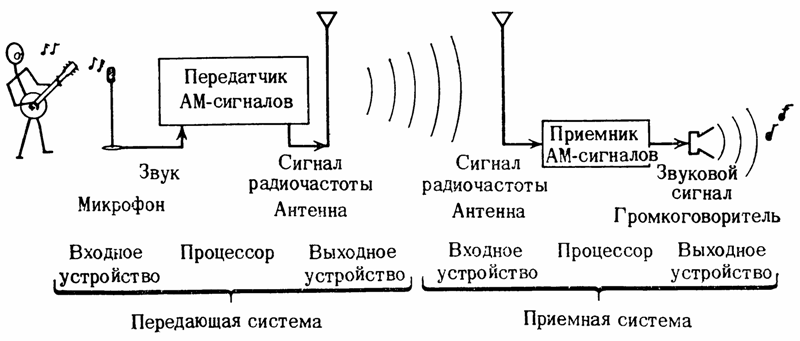 2016-04-20
2016-12-12
электрические сигналы, синусоидальный сигнал, прямоугольный сигнал, треугольный сигнал, пилообразный сигнал
2016-04-20
2016-12-12
электрические сигналы, синусоидальный сигнал, прямоугольный сигнал, треугольный сигнал, пилообразный сигнал
Первичные сигналы электросвязи | Основы электроакустики
Главная
Первичные сигналы электросвязи
Электрический сигнал это материальный носитель (переносчик) сообщения. В системах электросвязи для передачи сообщения на дальние расстояния переносчиком является переменный электрический ток (в проводных линиях), электромагнитное поле ( радиоволны), световые волны (оптоволоконнные линии связи). Скорость распространения перечисленных переносчиков приближается к скорости света и с помощью этих переносчиков можно передавать огромное количество информации.
В любой системе электросвязи должны быть устройства, осуществляющие преобразования:
- на передаче: информация — сообщение — сигнал;
- на приеме: сигнал — сообщение — информация.
Первичные сигналы электросвязи:
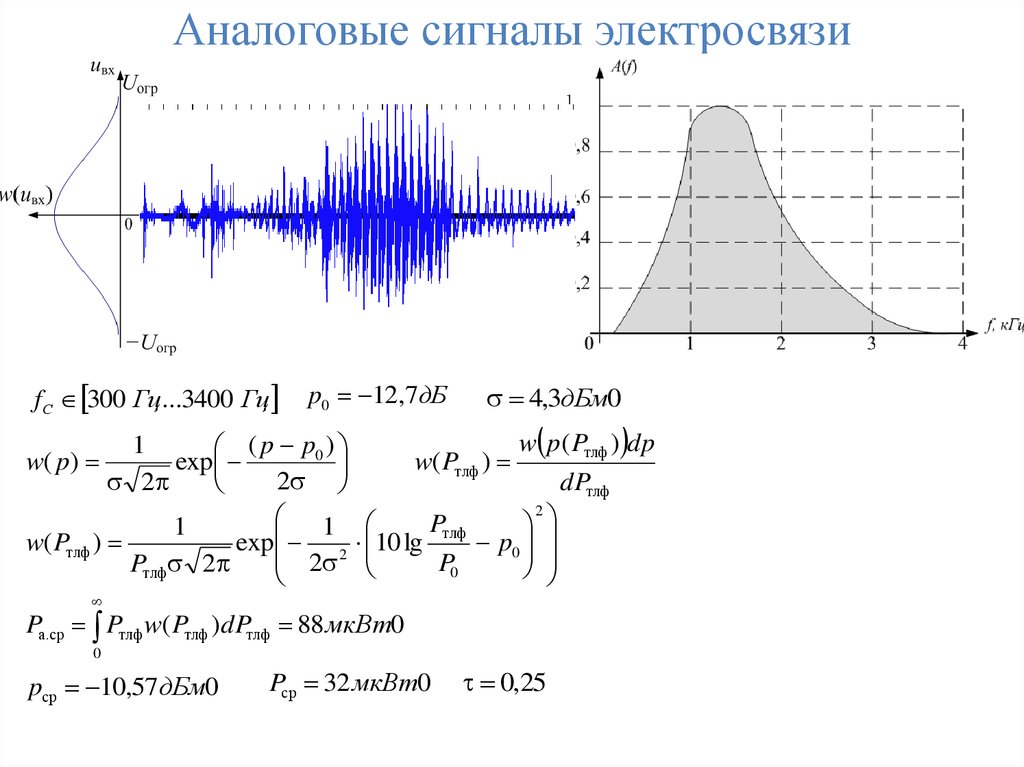
- телефонный речевой сигнал или сигнал ТЧ (тональной частоты), имеет спектр от 300 Гц до 3400 Гц; такой спектр достаточен для передачи речи по каналу связи и для принятия этого сигнала без искажений на приемной стороне;
- видеосигнал занимает полосу частот от 50Гц до 6,5 МГц (система SECAM) и от 50 Гц до 5,5 МГц (система PAL) – это самый широкополосный первичный сигнал;
- радиовещательный сигнал занимает полосу частот от 20 Гц до 20 кГц;
- Источник и получатель информации являются абонентами в системе связи, поэтому их называют абонентскими устройствами или терминалами.


Канал связи – это совокупность технических средств и среды распространения , обеспечивающих передачу сообщения от источника к получателю. В зависимости от вида сообщений и среды распространения различают каналы: телефонные, телеграфные, передачи данных, звукового и телевизионного вещания, проводные и кабельные, радиосвязи, цифровые. Математическая модель простого (гармонического) сигнала – это функция времени: u (t) = Um cos (2p f t + f), где Um – амплитуда, f – частота, f — фаза. Любой сложный сигнал (периодический и непериодический – импульсный на конечном отрезке времени) можно математически представить рядом Фурье: u (t) = Uo + Σ U mn cos (2πfn t + Φn), где Umn – амплитуда n–ой гармоники.
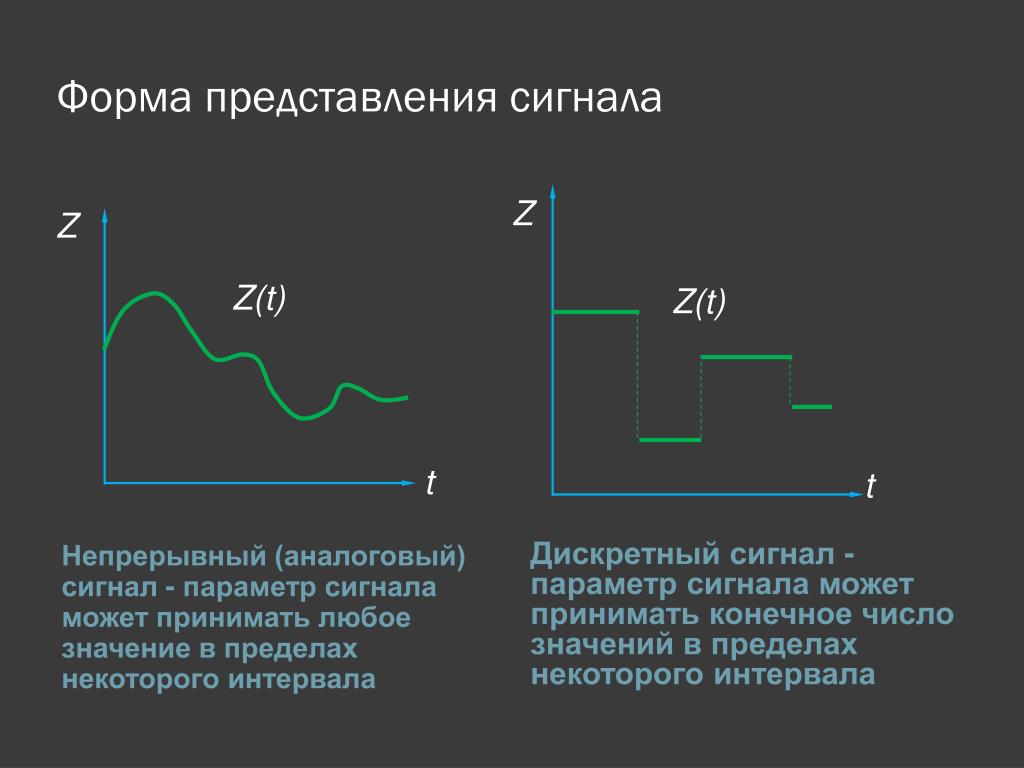 Различают непрерывные (аналоговые), дискретные и цифровые сигналы.
Различают непрерывные (аналоговые), дискретные и цифровые сигналы.Спектр сигнала
В настоящее время каналы электросвязи используются в основномдля передачи речевых, вещательных, телевизионных, факсимильных, телеграфных (передача данных) сигналов. Все они получили название первичных сигналов электросвязи.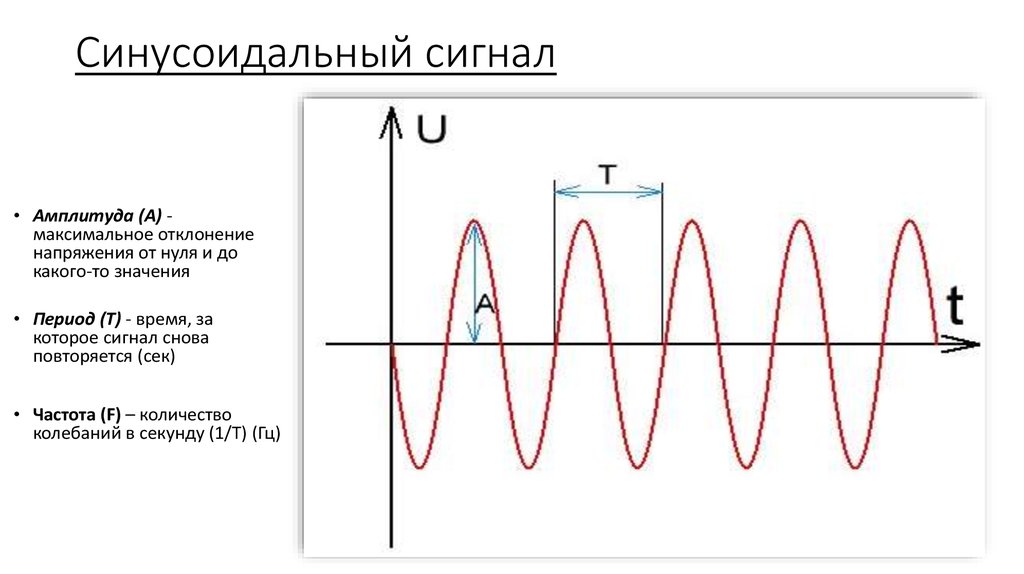
Речевой (телефонный) сигнал. Речь является случайным процессом. Первичные речевые сигналы, формируемые электроакустическими преобразователями — микрофонами, представляют собой реализации этого процесса. Статистические характеристики речевого сигнала получают усреднением результатов измерений как по множеству, так и по времени, При этом пользуются рекомендациями МККТТ. Абсолютный динамический уровень речевого сигнала L1 носит название волюм и измеряется специальным квадратичным вольтметром (волюмметром) с временем усреднения 200 мс. За нулевой уровень приняты Ро=1 мВт, Uo = 0,775 В при сопротивлении нагрузки 600 Ом. Одномерная плотность вероятности волюмов различных абонентов близка к нормальной. Спектральная плотность мощности речевого сигнала имеет максимум на частоте 300 …500 Гц и простирается от 50… 100 до 8000…. 10 000 Гц. Однако установлено, что качество речи вполне удовлетворительно при ограничении спектра частотами 300 … 3400 Гц. Качество восприятия речевого сигнала определяется разницей уровней сигнала и помех, поступающих на вход телефона.

Сигнал звукового вещания. Источниками звука при передаче вещательных программ обычно являются музыкальные инструменты или голос человека. Это также реализации случайного процесса. Для их качественного восприятия и полоса частот, и отношение сигнал-шум должны быть больше, чем для речевых сигналов. Характеристики псофометрического фильтра здесь несколько другие, псофометрический коэффициент — 6 дБ.
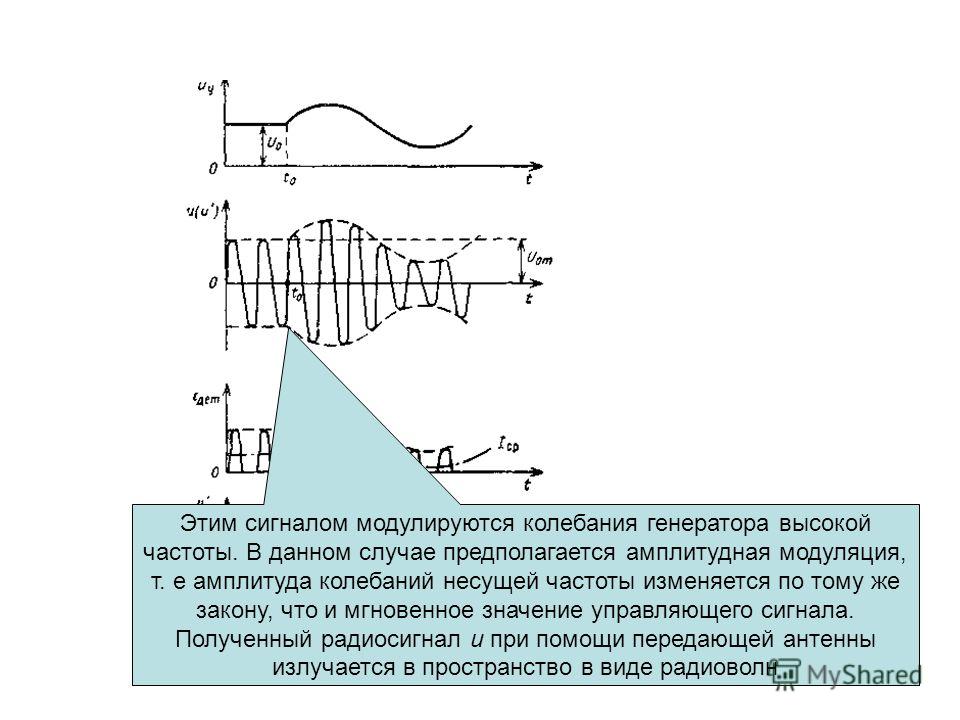
Телевизионный сигнал. В телевидении первичный сигнал формируется методом развертки. По принятому в СССР стандарту число строк 2 = 625. Передача движущегося изображения сводится к последовательной передаче кадров. В секунду передается 25 кадров, чтобы избежать мерцания на экране приемной трубки (кинескопа), стандартом предусмотрена чересстрочная развертка в виде двух полукадров. Первый полукадр — передача нечетных строк, второй — передача четных. Число передаваемых строк в секунду nz= 15625, время передачи одной строки — 64 мкс. Для синхронизации лучей приемной и передающей трубок необходима передача вспомогательных импульсов (синхронизирующих и гасящих обратный ход). Электрический сигнал, включающий в себя сигнал изображения и управляющие импульсы, называется полным телевизионным сигналом. Он состоит из случайного сигнала яркости и цветности (квазистационарный случайный процесс) и детерминированных вспомогательных импульсных сигналов. Помехи вызывают на экране кинескопа светлые или темные штрихи и пятна.
 При значительных помехах изображение видно как бы сквозь пелену мерцающих светлых и темных точек. На-блюдаются срывы кадровой и строчной синхронизации. Согласно рекомендациям МККТТ при изменениях мощности помехи в полосе телевизионного сигнала необходимо учитывать частотную чувствительность глаза. Для этого применяется специ-альный взвешивающий фильтр. На выходе такого фильтра уровень помехи на 9 дБ меньше, чем на его входе. Измерения проводятся для помехи с равномерным спектром. Защищенность телевизион-ного сигнала от взвешенной помехи должна быть не хуже 57 дБ.
При значительных помехах изображение видно как бы сквозь пелену мерцающих светлых и темных точек. На-блюдаются срывы кадровой и строчной синхронизации. Согласно рекомендациям МККТТ при изменениях мощности помехи в полосе телевизионного сигнала необходимо учитывать частотную чувствительность глаза. Для этого применяется специ-альный взвешивающий фильтр. На выходе такого фильтра уровень помехи на 9 дБ меньше, чем на его входе. Измерения проводятся для помехи с равномерным спектром. Защищенность телевизион-ного сигнала от взвешенной помехи должна быть не хуже 57 дБ.Факсимильный сигнал. Факсимильной связью называется передача неподвижных изображений (рисунков, чертежей, фотографий, текстов, газетных полос и т. д.) по каналам электросвязи. Первичные факсимильные сигналы получают в результате электрооптического анализа, заключающегося в преобразовании светового потока, отражаемого элементарными площадками изображения, в электрические сигналы. Как и в телевидении, здесь сигнал формируется методом развертки.
 Развертка обычно получается электромеханическим способом. Передающий факсимильный аппарат имеет барабан, на который накладывается изображение. На поверхность изображения проецируется яркое светлое пятно, перемещающееся вдоль оси барабана. Вращение барабана и смещение пятна обеспечивает развертку. Отраженный световой поток принимается фотоэлементом. В его цепи появляется ток, мгновенные значения которого определяются отражающей способностью элементов изображения. Характеристики тока зависят от размеров барабана, скорости его вращения, скорости смещения пятна по барабану, характера изображения. При передаче реальных изображений первичный сигнал получается сложной формы и его можно отнести к стационарному случайному процессу. В зависимости от характера изображения первичные факсимильные сигналы разделяются на штриховые, содержащие две градации яркости, и полутоновые со многими градациями яркости.
Развертка обычно получается электромеханическим способом. Передающий факсимильный аппарат имеет барабан, на который накладывается изображение. На поверхность изображения проецируется яркое светлое пятно, перемещающееся вдоль оси барабана. Вращение барабана и смещение пятна обеспечивает развертку. Отраженный световой поток принимается фотоэлементом. В его цепи появляется ток, мгновенные значения которого определяются отражающей способностью элементов изображения. Характеристики тока зависят от размеров барабана, скорости его вращения, скорости смещения пятна по барабану, характера изображения. При передаче реальных изображений первичный сигнал получается сложной формы и его можно отнести к стационарному случайному процессу. В зависимости от характера изображения первичные факсимильные сигналы разделяются на штриховые, содержащие две градации яркости, и полутоновые со многими градациями яркости. Телеграфный сигнал и сигнал передачи данных. Первичные телеграфные сигналы и сигналы передачи данных представляют собой случайную последовательность прямоугольных импульсов (однополярных или двуполярных) детерминированной амплитуды и длительности.
 Скорость модуляции, вероятности появления положительных(1) и отрицательных (0) импульсов, статистическая связь между ними определяется источником информации. Обычно Р(1) =Р(0) =0,5, т. е. импульсы равновероятны. Эффективную полосу частот последовательности телеграфных импульсов можно найти из соотношения Δf3≈1/tи=B Скорость модуляции В в современных системах телеграфии и передача данных достигает десятков тысяч бод.
Скорость модуляции, вероятности появления положительных(1) и отрицательных (0) импульсов, статистическая связь между ними определяется источником информации. Обычно Р(1) =Р(0) =0,5, т. е. импульсы равновероятны. Эффективную полосу частот последовательности телеграфных импульсов можно найти из соотношения Δf3≈1/tи=B Скорость модуляции В в современных системах телеграфии и передача данных достигает десятков тысяч бод.Формирование телевизионного сигнала
Шумы и помехи в трактах и каналах связи и вещания
Общие понятия о каналах и системах связи
O сохранении качества сигнала при цифровом преобразовании форматов
Цифровые системы многоканальной передачи информации
O сохранении качества сигнала при цифровой обработке
Методы построения многоканальных систем связи
Основные характеристики переменных электрических сигналов
Электротехника ТОЭ: 4.1. Классификация электрических сигналов
Теория / 4.1. Классификация электрических сигналов
Термин «сигнал» является распространенным в технике связи.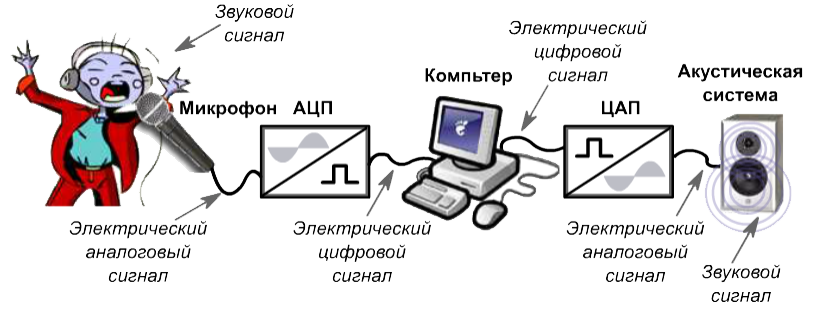 В соответствии с установившейся
традицией сигналом называют процесс изменения во времени физического состояния
какого-либо объекта.
В соответствии с установившейся
традицией сигналом называют процесс изменения во времени физического состояния
какого-либо объекта.
Электрический сигнал – это электрическое напряжение или ток, изменяющиеся с течением времени и несущие в себе информацию.
Сигналы различаются по частоте: высокочастотные и низкочастотные сигналы. Высокочастотный сигнал может быть промодулирован низкой частотой, такой сигнал называется модулированным сигналом (рис. 4.1, а). В отсутствие модуляции – немодулированный ВЧ сигнал (рис. 4.1, б). Высокочастотная составляющая модулированного сигнала называется несущей частотой.
Сигналы подразделяются на периодические и непериодические. У периодических сигналов форма сигнала повторяется через определенный промежуток времени, у непериодических эта особенность отсутствует.
Периодические сигналы принято подразделять на гармонические и импульсные.
Гармонические – это сигналы, изменяющиеся по закону синусов.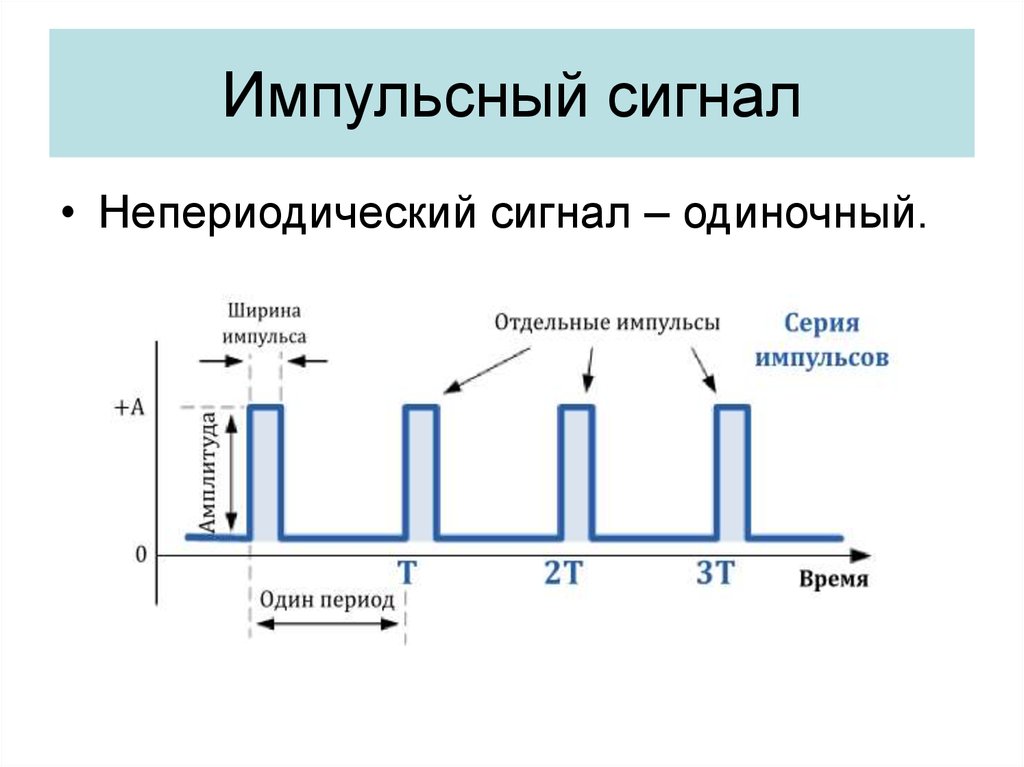
Импульсные сигналы – это колебания, присутствующие лишь в пределах конечного отрезка времени. При этом различают видеоимпульсы и радиоимпульсы. Радиоимпульсы представляют собой модулированные высокочастотные сигналы – волновые пакеты, видеоимпульсы – импульсы напряжения (тока), действующие в течение коротких промежутков времени, меньших по сравнению с длительностью переходных процессов.
Импульсы могут иметь различную форму: прямоугольную, трапецеидальную, треугольную, колоколообразную.
Периодическая последовательность импульсов характеризуется периодом следования импульсов T0 и частотой следования импульсов fи = 1 / Tи, длительностью
импульса Ти длительностью паузы между импульсами Tп , скважностью, то есть
отношением периода следования импульсов к длительности импульса Q = T0 / Tи, коэффициентом заполнения, то есть отношением периода следования
импульсов к длительности паузы Kз = T0 / Tп .
Сигналы бывают аналоговые и цифровые.
Аналоговый сигнал может принимать любое произвольное значение напряжения или тока в заданном допустимом диапазоне от минимального значения до максимального значения. То есть физический процесс, порождающий сигнал, развивается во времени таким образом, что значения сигнала можно измерять в любые моменты времени. С аналоговыми сигналами работают аналоговые вычислительные машины и устройства, аналоговый сигнал часто формируется и на выходах датчиков информации.
Первоначально в радиотехнике использовались сигналы исключительно аналогового типа. Такие сигналы позволяли с успехом решать относительно несложные технические задачи (радиосвязь, телевидение). Аналоговые сигналы было просто генерировать, принимать и обрабатывать с помощью доступных средств.
Возросшие требования к радиотехническим
системам, разнообразие применения заставили искать новые принципы их
построения. На смену аналоговым в ряде случаев пришли импульсные системы,
работа которых основана на использовании дискретных сигналов.
Простейшая математическая модель дискретного сигнала sd(t) – это счетное множество точек {ti} (i – целое число) на оси времени, в каждой из которых определено отсчетное значение сигнала si. Как правило, шаг дискретизации для каждого сигнала постоянен.
Одно из преимуществ дискретных сигналов по сравнению с аналоговыми – отсутствие необходимости воспроизводить сигнал непрерывно во все моменты времени. За счет этого появляется возможность по одной и той же линии связи передавать сообщения от разных источников, организуя многоканальную связь с разделением каналов во времени.
Особой разновидностью дискретных сигналов являются цифровые сигналы. Для них характерно то, что отсчетные значения представлены в форме чисел. Из соображений технического удобства реализации и обработки обычно используют двоичную систему чисел.
Следует отметить, что любой дискретный или
цифровой сигнал по сути является сигналом аналоговым. Так, медленно меняющемуся
во времени аналоговому сигналу s(t),
можно поставить в соответствие его дискретный образ, имеющий вид
последовательности прямоугольных видеоимпульсов одинаковой длительности с
высотой, пропорциональной значениям s(t)
в отсчетных точках, либо с постоянной высотой, но различной длительностью в
отсчетных точках (рис. 4.2).
Так, медленно меняющемуся
во времени аналоговому сигналу s(t),
можно поставить в соответствие его дискретный образ, имеющий вид
последовательности прямоугольных видеоимпульсов одинаковой длительности с
высотой, пропорциональной значениям s(t)
в отсчетных точках, либо с постоянной высотой, но различной длительностью в
отсчетных точках (рис. 4.2).
Оба способа становятся эквивалентными, если положить, что значения аналогового сигнала в точках дискретизации пропорциональны площади отдельных видеоимпульсов.
Как аналоговый сигнал преобразуется в цифровой
Телевизионный сигнал — совокупность электрических сигналов, содержащая информацию о телевизионном изображении и звуке. Телевизионный сигнал может передаваться по радио или по кабелю. Термин употребляется в большинстве случаев применительно к аналоговому телевидению, потому что цифровое оперирует таким понятием, как поток данных.
В электронике сигналы делят на: аналоговые, дискретные и цифровые. Начнем с того, что все, что мы чувствуем, видим, слышим в большинстве своем является аналоговым сигналом, а то, что видит процессор компьютера – это цифровой сигнал. Звучит не совсем понятно, поэтому давайте разбираться с этими определениями и с тем как один вид сигналов преобразовывается в другой.
Начнем с того, что все, что мы чувствуем, видим, слышим в большинстве своем является аналоговым сигналом, а то, что видит процессор компьютера – это цифровой сигнал. Звучит не совсем понятно, поэтому давайте разбираться с этими определениями и с тем как один вид сигналов преобразовывается в другой.
Типы сигналов
В электрическом представлении аналоговый сигнал, судя по его названию, является аналогом реальной величины. Например, вы чувствуете температуру окружающей среды постоянно, на протяжении всей жизни. Нет никаких перерывов. При этом вы чувствуете не только два уровня «горячо» и «холодно», а бесконечное число ощущений, которые описывают эту величину.
Для человека «холодно» может быть по разному, это и осенняя прохлада и зимний мороз, и легкие заморозки, но не всегда «холодно» это отрицательная температура, как и «тепло» — не всегда положительная температура.
Отсюда следует, что у аналогового сигнала две особенности:
1. Непрерывность во времени.
2. Число величин сигнала стремится к бесконечности, т.е. аналоговый сигнал нельзя точно поделить на части или проградуировать, разбив шкалу на конкретные участки. Способы измерения – основаны на единице измерений, и их точность зависит лишь от цены деления шкалы, чем она меньше, тем точнее измерение.
Дискретные сигналы – это сигналы, которые представляют собой последовательность отчетов или измерений какой-либо величины. Измерения таких сигналов не непрерывны, а периодичны.
Попытаюсь объяснить. Если вы установили термометр где-нибудь он измеряет аналоговую величину – это следует из вышеописанного. Но вы, фактически следя за его показаниями, получаете дискретную информацию. Дискретный – значит отдельный.
Например, вы проснулись и узнали, сколько градусов на термометре, в следующий раз вы на него посмотрели на градусник в полдень, и третий раз вечером. Вы не знаете, с какой скоростью изменялась температура, равномерно, или резким скачком, вы знаете только данные в тот момент времени, который наблюдали.
Цифровые сигналы – это набор уровней, типа 1 и 0, высокий и низкий, есть или нет. Глубина отражения информации в цифровом виде ограничена разрядностью цифрового устройства (набора логики, микроконтроллера, процессора etc.) Получается что для хранения булевых данных он подходит идеально. Пример, можно привести следующий, для хранений данных типа «День» и «Ночь», достаточно 1 бита информации.
Бит – это минимальная величина представления информации в цифровом виде, в нём может храниться только два типа значений 1 (логическая единица, высокий уровень), или 0 (логический ноль, низкий уровень).
В электронике бит информации представляется в виде низкого уровня напряжения (близкое к 0) и высокого уровня напряжения (зависит от конкретного устройства, часто совпадает с напряжением питания данного цифрового узла, типовые значения – 1.7, 3.3. 5В, 15В).
Все промежуточные значения между принятыми низким и высоким уровнем являются переходной областью и могут не обладать конкретным значением, в зависимости от схемотехники, как устройства в целом, так и внутренней схемы микроконтроллера (или любого другого цифрового устройства) могут иметь разный переходный уровень, например для 5-тивольтовой логики за ноль могут приниматься значения напряжения от 0 до 0.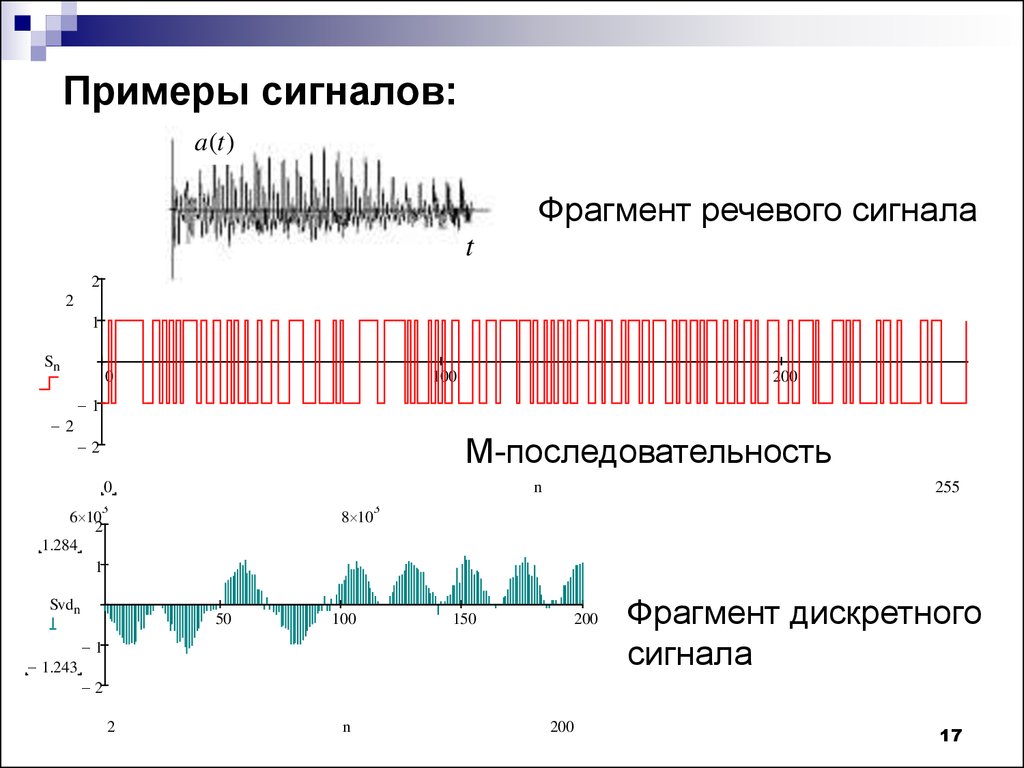 8В, а за единицу от 2В до 5В, при этом промежуток между 0.8 и 2В – это неопределенная зона, фактически с ее помощью отделяется ноль от единицы.
8В, а за единицу от 2В до 5В, при этом промежуток между 0.8 и 2В – это неопределенная зона, фактически с ее помощью отделяется ноль от единицы.
Чем более точные и ёмкие значения нужно хранить, тем больше нужно бит, приведем таблицу-пример с отображением в цифровом виде четырёх значений времени суток:
Ночь – Утро – День – Вечер
Для этого нам нужно уже 2 бита:
Аналогово-цифровое преобразование
В общем случае аналогово-цифровым преобразованием называется процесс перевода физической величины в цифровое значение. Цифровым значением является набор единиц и нолей воспринятых обрабатывающим устройством.
Такое преобразование нужно для взаимодействия цифровой техники с окружающей средой.
Так как аналоговый электрический сигнал повторяет своей формой входной сигнал, он не может быть записан в цифровом виде «так как есть» поскольку он имеет бесконечное число значений. Примером можно привести процесс записи звука. Он в первичном виде выглядит так:
Он представляет собой сумму волн с различными частотами. Которые, при разложении по частотам (подробнее об этом смотрите преобразования Фурье), так или иначе, можно приблизить к похожей картинке:
Которые, при разложении по частотам (подробнее об этом смотрите преобразования Фурье), так или иначе, можно приблизить к похожей картинке:
Теперь попробуйте это представить в виде набора типа «111100101010100», довольно сложно, не так ли?
Другим примером необходимости преобразования аналоговой величины в цифровую, является её измерение: электронные термометры, вольтметры, амперметры и прочие измерительные приборы взаимодействую с аналоговыми величинами.
Как происходит преобразование?
Сначала посмотрите на схему типового преобразования аналогового сигнала в цифровой и обратно. Позже мы к ней вернемся.
Фактически это сложный процесс, который состоит из двух основных этапов:
1. Дискретизация сигнала.
2. Квантование по уровню.
Дискретизация сигнала это определения промежутков времени, на которых измеряется сигнал. Чем короче эти промежутки – тем точнее измерение. Периодом дискретизации (Т) называется отрезок времени от начала считывания данных до его конца. Частота дискретизации (f) – это обратная величина:
Частота дискретизации (f) – это обратная величина:
fд=1/Т
После считывания сигнала происходит его обработка и сохранение в память.
Получается, что за время, которое считываются и обрабатываются показания сигнала, он может измениться, таким образом, происходит искажение измеряемой величины. Есть такая теорема Котельникова и из нее вытекает такое правило:
Частота дискретизации должны быть как минимум в 2 раза больше чем частота дискретизируемого сигнала.
Это скриншот из википедии, с выдержкой из теоремы.
Для определения численного значение необходимо квантование по уровню. Квант – это определенный промежуток измеряемых значений, усреднено приведенный к определенному числу.
X1…X2=Xy
Т.е. сигналы величиной от X1 до X2, условно приравнивается к определенному значению Xy. Это напоминает цену деления стрелочного измерительного прибора. Когда вы снимаете показания, зачастую вы их равняете по ближайшей отметке на шкале прибора.
Так и с квантованием по уровню, чем больше квантов, тем более точные измерения и тем больше знаков после запятой (сотых, тысячных и так далее значений) они могут содержать.
Точнее сказать число знаков после запятой скорее определяется разрядностью АЦП.
На картинке изображен процесс квантования сигнала с помощью одного бита информации, как я описывал выше, когда при превышении определенного предела принимается значение высокого уровня.
Справа показано квантование сигнала, и запись в виде двух бит данных. Как видите, этот фрагмент сигнала разбит уже на четыре значения. Получается, что в результате плавный аналоговый сигнал превратился в цифровой «ступенчатый» сигнал.
Количество уровней квантования определяется по формуле:
де n — количество разрядов, N — уровень квантования.
Вот пример сигнала разбитого на большее число квантов:
Отсюда очень хорошо видно, что чем чаще снимаются значения сигнала (больше частота дискретизации), тем точнее он измеряется.
На этой картинке изображено преобразование аналогового сигнала в цифровой вид, а слева от оси ординат (вертикальной оси) запись в цифровом 8-битном виде.
Аналогово-цифровые преобразователи
АЦП или Аналогово-цифровой преобразователь может выполняться в виде отдельного устройства или быть встроенным в микроконтроллер.
Ранее в микроконтроллеры, например семейства MCS-51, не содержали в своем составе АЦП, использовалась для этого внешняя микросхема и возникала необходимость писать подпрограмму обработки значений внешней ИМС.
Сейчас они есть в большинстве современных микроконтроллеров, например AVR AtMEGA328, который является основой большинства популярных плат Ардуино, он встроен в сам МК. На языке Arduino чтение аналоговых данных осуществляется просто – командой AnalogRead(). Хотя в микропроцессоре, который установлен в той же не менее популярной Raspberry PI его нет, так что не все так однозначно.
Фактически существует большое число вариантов аналогово-цифровых преобразователей, у каждого из которых есть свои недостатки и преимущества. Описывать которые в пределах этой статьи не имеет особого смысла, так как это большой объём материала.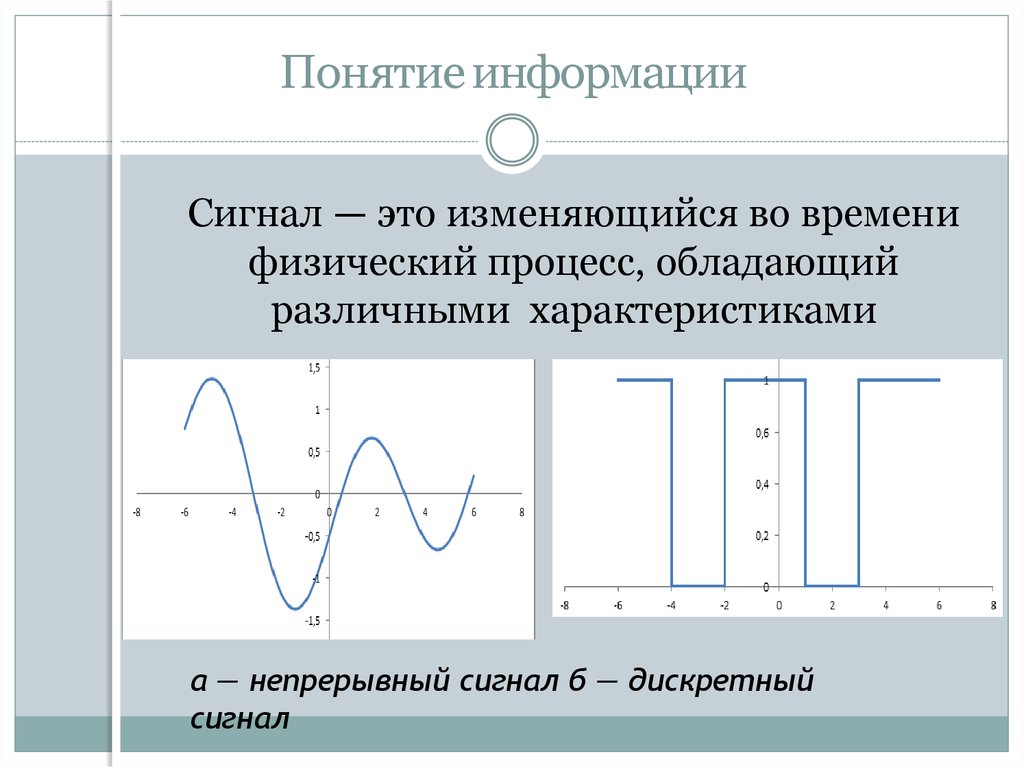 Рассмотрим лишь общую структуру некоторых из них.
Рассмотрим лишь общую структуру некоторых из них.
Самым старым запатентованным вариантом АЦП, является патент Paul M. Rainey, «Facsimile Telegraph System,» U.S. Patent 1,608,527, Filed July 20, 1921, Issued November 30, 1926. Это 5-ти битный АЦП прямого преобразования. Из названия патента приходят мысли о том, что использование этого прибора было связано с передачей данных через телеграф.
Если говорить о современных АЦП прямого преобразования имеют следующую схему:
Отсюда видно, что вход представляет собой цепочку из компараторов, которые на выходе своем выдают сигнал при пересечении какого-то порогового сигнала. Это и есть разрядность и квантование. Кто хоть немного силен в схемотехнике, увидел этот очевидный факт.
Кто не силен, то входная цепь работает таким образом:
Аналоговый сигнал поступает на вход «+», на все сразу. На выходы с обозначением «-» поступает опорное напряжение, которое раскладывается с помощью цепочки резисторов (резистивного делителя) на ряд опорных напряжений.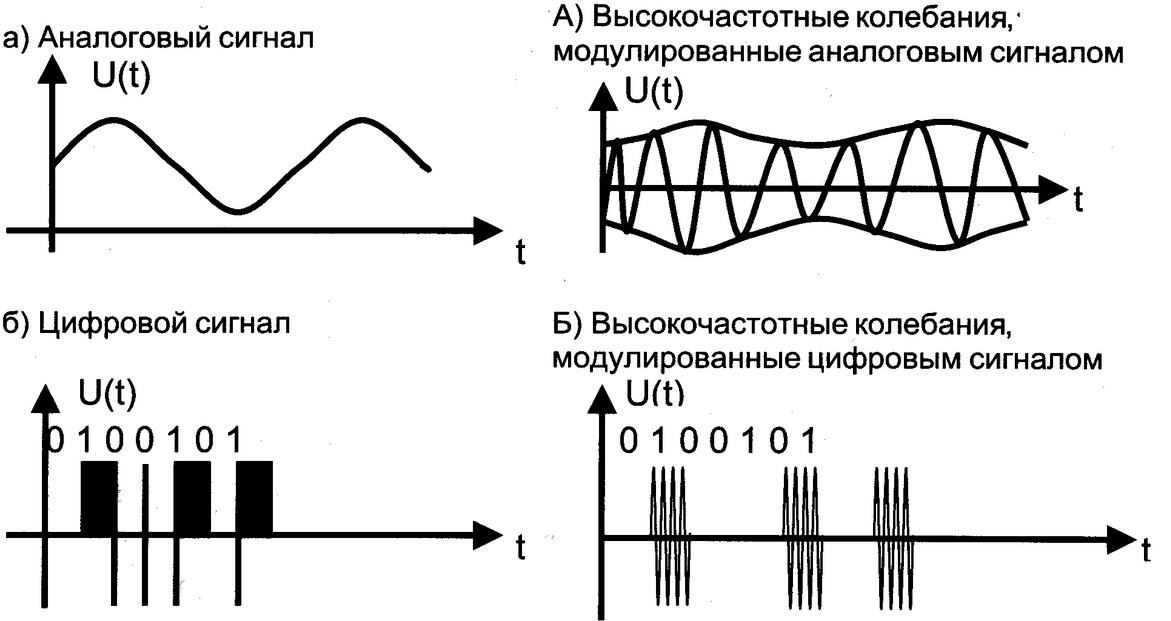 К примеру, ряд для этой цепи выглядит наподобие такого соотношения:
К примеру, ряд для этой цепи выглядит наподобие такого соотношения:
Urefi=(1/16, 3/16, 5/16, 7/16, 9/16, 11/16, 13/16)*Uref
В скобках через запятую указано, какую часть от общего опорного напряжения Uref подают на вход каждого входного напряжения.
Т.е. каждый из элементов имеет два входа, когда напряжение на входе со знаком «+»превышает напряжение на входе со знаком «-», то на его выходе появляется логическая единица. Когда на положительном (неинвертирующем) входе напряжение меньше, чем на отрицательно (инвертирующем), то на выходе – ноль.
Напряжение делиться таким образом, чтобы входное напряжение разбить на нужное количество разрядов. При достижении напряжения на входе на выходе соответствующего элемента появляется сигнал, схема обработки выводит «правильный» сигнал в цифровом виде.
Такой компаратор хорош скоростью обработки данных, все элементы входной цепи срабатывают параллельно, основная задержка этого вида АЦП формируется из задержки 1 компаратора (все же одновременно параллельно срабатывают) и задержки шифратор. 8 компараторов, а это целых 256 штук. Для десятиразрядного (в ардуино 10-разрядный АЦП, кстати, но другого типа) нужно 1024 компаратора. Судите сами о целесообразности такого варианта обработки, и где он может понадобиться.
8 компараторов, а это целых 256 штук. Для десятиразрядного (в ардуино 10-разрядный АЦП, кстати, но другого типа) нужно 1024 компаратора. Судите сами о целесообразности такого варианта обработки, и где он может понадобиться.
Есть и другие виды АЦП:
Заключение
Преобразование аналогового сигнала в цифровой нужно для считывания параметров с аналоговых датчиков. Есть отдельный вид цифровых датчиков, они представляют собой либо интегральные микросхемы, например DS18b20 – на его выходе уже цифровой сигнал и его можно обрабатывать любыми микроконтроллерами или микропроцессорами без необходимости применения АЦП, или аналоговый датчик на плате на которой уже размещен свой преобразователь. У каждого типа датчиков есть свои плюсы и минусы, такие как помехоустойчивость и погрешность измерений.
Знание принципов преобразование обязательно для всех кто работает с микроконтроллерами, ведь не в каждой даже современной системе встроены такие преобразователи, приходится использовать внешние микросхемы. Для примера можно привести такую плату, разработанную специально под GPIO-разъём Raspberry PI, с прецизионным АЦП на ADS1256.
Для примера можно привести такую плату, разработанную специально под GPIO-разъём Raspberry PI, с прецизионным АЦП на ADS1256.
Ранее ЭлектроВести писали, что власти Вены запустили тестирование технологии умных камер, которые будут распознавать приближение пешеходов и подавать сигнал на светофор.
По материалам: electrik.info.
Аналоговый электрический сигнал это
Содержание:
- 0.0.1 Типы сигналов
- 0.0.1.1 Обработка сигнала
- 0.0.1.2 Аналоговый сигнал
- 0.0.1.3 Цифровой сигнал
- 0.0.1.4 Аналоговый и цифровой сигнал и цифро-аналоговое преобразования
- 0.0.1.4.1 Что происходит в мобильных телефонах, которые передают информацию по цифровым каналам
- 0.0.2 Типы сигналов
- 0.0.2.1 Обработка сигнала
- 0.0.2.2 Аналоговый сигнал
- 0.0.2.3 Цифровой сигнал
- 0.0.2.4 Аналоговый и цифровой сигнал и цифро-аналоговое преобразования
- 0.0.2.4.1 Что происходит в мобильных телефонах, которые передают информацию по цифровым каналам
- 0.0.1 Типы сигналов
- 1 Сигнал
- 2 Виды сигналов
- 3 Для чего обрабатывается сигнал?
- 4 Создание и формирование
- 5 Динамический диапазон
- 6 Аналоговый сигнал
- 7 Цифровой сигнал
- 8 Применение цифрового сигнала
- 9 Дискретный сигнал
- 10 Сравнение цифрового и аналогового сигналов
Сигналами называют информационные коды, которые применяются людьми для того, чтобы передавать сообщения в информационной системе. Сигнал может подаваться, но его получение не обязательно. Тогда как сообщением можно считать только такой сигнал (или совокупность сигналов), который был принят и декодирован получателем (аналоговый и цифровой сигнал).
Сигнал может подаваться, но его получение не обязательно. Тогда как сообщением можно считать только такой сигнал (или совокупность сигналов), который был принят и декодирован получателем (аналоговый и цифровой сигнал).
Одними из первых методов передачи информации без участия людей или других живых существ были сигнальные костры. При возникновении опасности последовательно разводились костры от одного поста к другому. Далее мы будем рассматривать способ передачи информации при помощи электромагнитных сигналов и подробно остановимся на рассмотрении темы аналоговый и цифровой сигнал .
Любой сигнал может быть представлен в виде функции, которая описывает изменения его характеристик. Такое представление удобно для изучения устройств и систем радиотехники. Помимо сигнала в радиотехнике есть еще шум, который является его альтернативой. Шум не несет полезной информации и искажает сигнал, взаимодействуя с ним.
Само понятие дает возможность отвлечься от конкретных физических величин при рассмотрении явлений, связанных с кодированием и декодированием информации. Математическая модель сигнала в исследованиях позволяет опираться на параметры функции времени.
Математическая модель сигнала в исследованиях позволяет опираться на параметры функции времени.
Сигналы по физической среде носителя информации делятся на электрические, оптические, акустические и электромагнитные.
По методу задания сигнал может быть регулярным и нерегулярным. Регулярный сигнал представляется детерминированной функцией времени. Нерегулярный сигнал в радиотехнике представлен хаотической функцией времени и анализируется вероятностным подходом.
Сигналы в зависимости от функции, которая описывает их параметры могут быть аналоговыми и дискретными. Дискретный сигнал, который был подвергнут квантованию называется цифровым сигналом.
Обработка сигналаАналоговый и цифровой сигнал обрабатывается и направлен на то, чтобы передать и получить информацию, закодированную в сигнале. После извлечения информации ее можно применять в разных целях. В частных случаях информация подвергается форматированию.
Аналоговые сигналы подвергаются усилению, фильтрации, модуляции и демодуляции. Цифровые же помимо этого еще могут подвергаться сжатию, обнаружению и др.
Аналоговый сигналНаши органы чувств воспринимают всю поступающую в них информацию в аналоговом виде. К примеру, если мы видим проезжающий мимо автомобиль, мы видим его движение непрерывно. Если бы наш мозг мог получать информацию о его положении раз в 10 секунд, люди бы постоянно попадали под колеса. Но мы можем оценивать расстояние куда быстрее и это расстояние в каждый момент времени четко определено.
Абсолютно то же самое происходит и с другой информацией, мы можем оценивать громкость в любой момент, чувствовать какое давление наши пальцы оказывают на предметы и т.п. Иными словами, практически вся информация, которая может возникать в природе имеет аналоговый вид. Передавать подобную информацию проще всего аналоговыми сигналами, которые являются непрерывными и определены в любой момент времени.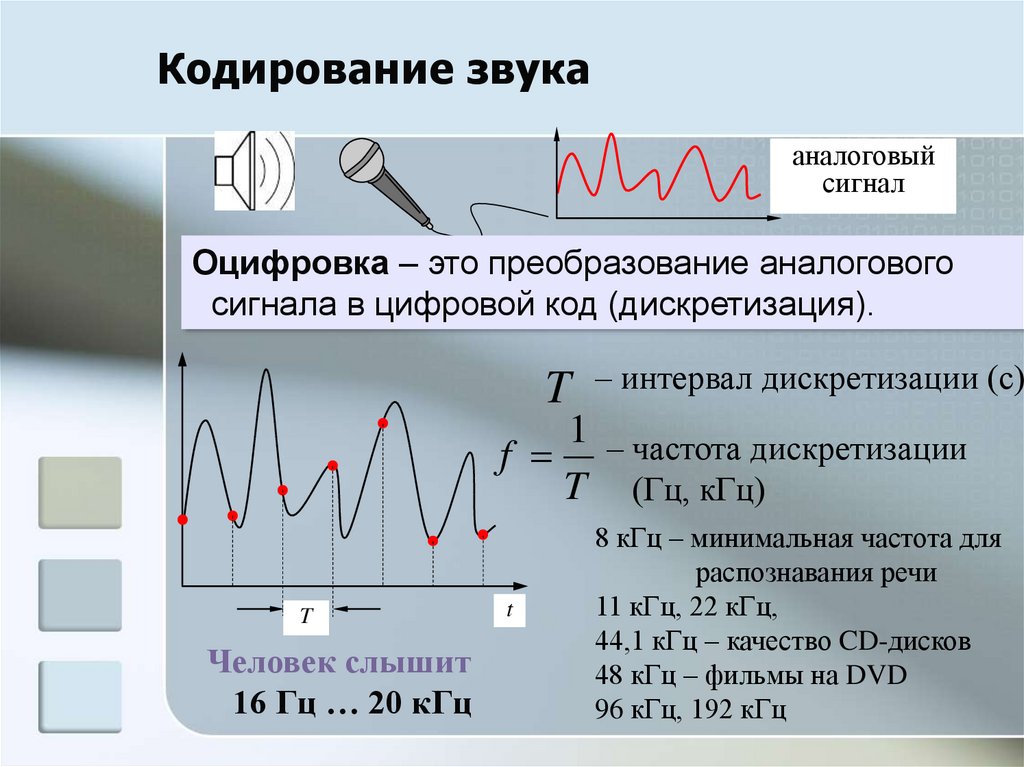
Чтобы понять, как выглядит аналоговый электрический сигнал, можно представить себе график, на котором будет отображена амплитуда по вертикальной оси и время по горизонтальной оси. Если мы, к примеру, замеряем изменение температуры, то на графике появится непрерывная линия, отображающая ее значение в каждый момент времени. Чтобы передать такой сигнал с помощью электрического тока, нам надо сопоставить значение температуры со значением напряжения. Так, например, 35.342 градуса по Цельсию могут быть закодированы как напряжение 3.5342 В.
Аналоговые сигналы раньше использовались во всех видах связи. Чтобы избежать помех такой сигнал нужно усиливать. Чем выше уровень шума, то есть помех, тем сильнее надо усиливать сигнал, чтобы его можно было принять без искажения. Такой метод обработки сигнала затрачивает много энергии на выделение тепла. При этом усиленный сигнал может сам стать причиной помех для других каналов связи.
Сейчас аналоговые сигналы еще применяются в телевидении и радио, для преобразования входного сигнала в микрофонах. Но, в целом, этот тип сигнала повсеместно вытеснен или вытесняется цифровыми сигналами.
Но, в целом, этот тип сигнала повсеместно вытеснен или вытесняется цифровыми сигналами.
Цифровой сигнал представлен последовательностью цифровых значений. Чаще всего сейчас применяются двоичные цифровые сигналы, так как они используются в двоичной электронике и легче кодируются.
В отличие от предыдущего типа сигнала цифровой сигнал имеет два значения «1» и «0». Если мы вспомним наш пример с измерением температуры, то тут сигнал будет сформирован иначе. Если напряжение, которое подается аналоговым сигналом соответствует значению измеряемой температуры, то в цифровом сигнале для каждого значения температуры будет подаваться определенное количество импульсов напряжения. Сам импульс напряжения тут будет равен «1», а отсутствие напряжения – «0». Приемная аппаратура будет декодировать импульсы и восстановит исходные данные.
Представив, как будет выглядеть цифровой сигнал на графике, мы увидим, что переход от нулевого значения к максимальному производится резко. Именно эта особенность позволяет принимающей аппаратуре более четко «видеть» сигнал. Если возникают какие-либо помехи, приемнику проще декодировать сигнал, нежели чем при аналоговой передаче.
Именно эта особенность позволяет принимающей аппаратуре более четко «видеть» сигнал. Если возникают какие-либо помехи, приемнику проще декодировать сигнал, нежели чем при аналоговой передаче.
Однако цифровой сигнал с очень большим уровнем шума восстановить невозможно, тогда как из аналогового типа при большом искажении еще есть возможность «выудить» информацию. Это связано с эффектом обрыва. Суть эффекта в том, что цифровые сигналы могут передаваться на определенные расстояния, а затем просто обрываются. Этот эффект возникает повсеместно и решается простой регенерацией сигнала. Там, где сигнал обрывается, нужно вставить повторитель или уменьшить длину линии связи. Повторитель не усиливает сигнал, а распознает его изначальный вид и выдает его точную копию и может использоваться сколь угодно в цепи. Такие способы повторения сигнала активно применяются в сетевых технологиях.
Помимо всего прочего аналоговый и цифровой сигнал различается и возможность кодирования и шифрования информации.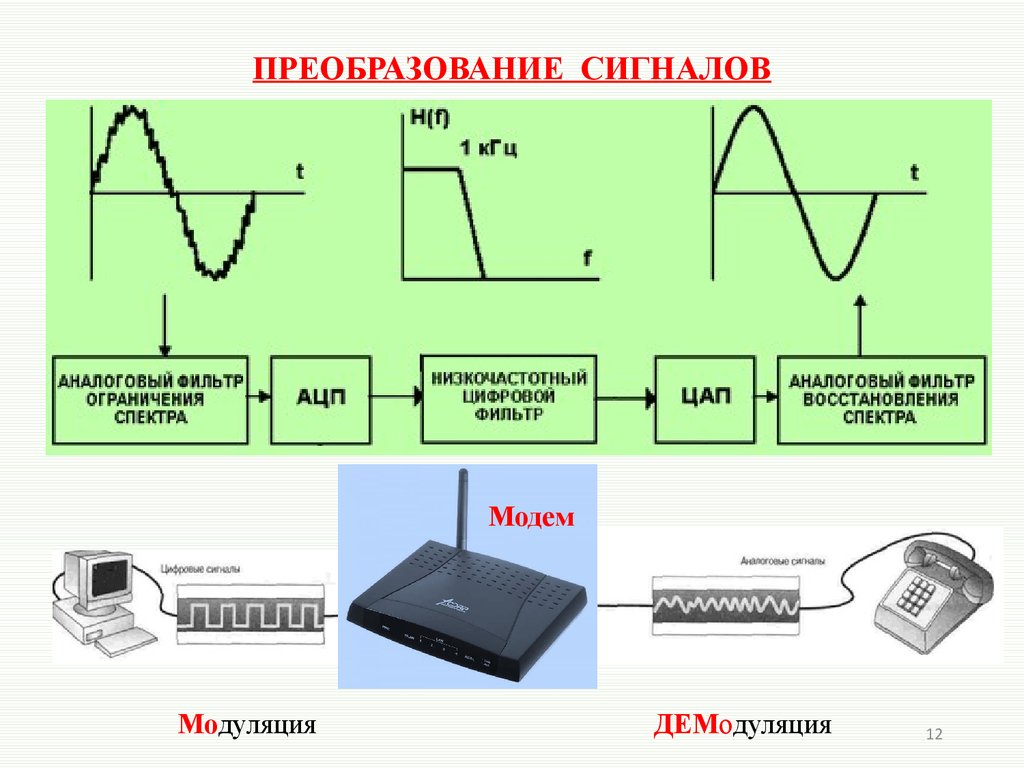 Это является одной из причин перехода мобильной связи на «цифру».
Это является одной из причин перехода мобильной связи на «цифру».
Следует еще немного рассказать о том, как аналоговая информация передается по цифровым каналам связи. Вновь прибегнем к примерам. Как уже говорилось звук – это аналоговый сигнал.
Что происходит в мобильных телефонах, которые передают информацию по цифровым каналам
Звук, попадая в микрофон подвергается аналого-цифровому преобразованию (АЦП). Этот процесс состоит из 3 ступеней. Берутся отдельные значения сигнала через одинаковые отрезки времени, этот процесс называется дискретизация. По теореме Котельникова о пропускной способности каналов, частота взятия этих значений должна быть вдвое выше, чем самая высокая частота сигнала. То есть, если в нашем канале стоит ограничение на частоту в 4 кГц, то частота дискретизации будет составлять 8 кГц.
Далее все выбранные значения сигнала округляются или, иначе говоря, квантуются. Чем больше уровней при этом будет создано, тем выше будет точность восстановленного сигнала на приемнике. Затем все значения преобразуются в двоичный код, который передается на базовую станцию и затем доходит до другого абонента, являющегося приемником. В телефоне приемника происходит процедура цифро-аналогового преобразования (ЦАП). Это обратная процедура, цель которой на выходе получить сигнал как можно более идентичный исходному. Далее уже аналоговый сигнал выходит в виде звука из динамика телефона.
Чем больше уровней при этом будет создано, тем выше будет точность восстановленного сигнала на приемнике. Затем все значения преобразуются в двоичный код, который передается на базовую станцию и затем доходит до другого абонента, являющегося приемником. В телефоне приемника происходит процедура цифро-аналогового преобразования (ЦАП). Это обратная процедура, цель которой на выходе получить сигнал как можно более идентичный исходному. Далее уже аналоговый сигнал выходит в виде звука из динамика телефона.
Сигналами называют информационные коды, которые применяются людьми для того, чтобы передавать сообщения в информационной системе. Сигнал может подаваться, но его получение не обязательно. Тогда как сообщением можно считать только такой сигнал (или совокупность сигналов), который был принят и декодирован получателем (аналоговый и цифровой сигнал).
Одними из первых методов передачи информации без участия людей или других живых существ были сигнальные костры. При возникновении опасности последовательно разводились костры от одного поста к другому. Далее мы будем рассматривать способ передачи информации при помощи электромагнитных сигналов и подробно остановимся на рассмотрении темы аналоговый и цифровой сигнал .
Далее мы будем рассматривать способ передачи информации при помощи электромагнитных сигналов и подробно остановимся на рассмотрении темы аналоговый и цифровой сигнал .
Любой сигнал может быть представлен в виде функции, которая описывает изменения его характеристик. Такое представление удобно для изучения устройств и систем радиотехники. Помимо сигнала в радиотехнике есть еще шум, который является его альтернативой. Шум не несет полезной информации и искажает сигнал, взаимодействуя с ним.
Само понятие дает возможность отвлечься от конкретных физических величин при рассмотрении явлений, связанных с кодированием и декодированием информации. Математическая модель сигнала в исследованиях позволяет опираться на параметры функции времени.
Типы сигналовСигналы по физической среде носителя информации делятся на электрические, оптические, акустические и электромагнитные.
По методу задания сигнал может быть регулярным и нерегулярным. Регулярный сигнал представляется детерминированной функцией времени. Нерегулярный сигнал в радиотехнике представлен хаотической функцией времени и анализируется вероятностным подходом.
Нерегулярный сигнал в радиотехнике представлен хаотической функцией времени и анализируется вероятностным подходом.
Сигналы в зависимости от функции, которая описывает их параметры могут быть аналоговыми и дискретными. Дискретный сигнал, который был подвергнут квантованию называется цифровым сигналом.
Обработка сигналаАналоговый и цифровой сигнал обрабатывается и направлен на то, чтобы передать и получить информацию, закодированную в сигнале. После извлечения информации ее можно применять в разных целях. В частных случаях информация подвергается форматированию.
Аналоговые сигналы подвергаются усилению, фильтрации, модуляции и демодуляции. Цифровые же помимо этого еще могут подвергаться сжатию, обнаружению и др.
Аналоговый сигналНаши органы чувств воспринимают всю поступающую в них информацию в аналоговом виде. К примеру, если мы видим проезжающий мимо автомобиль, мы видим его движение непрерывно. Если бы наш мозг мог получать информацию о его положении раз в 10 секунд, люди бы постоянно попадали под колеса.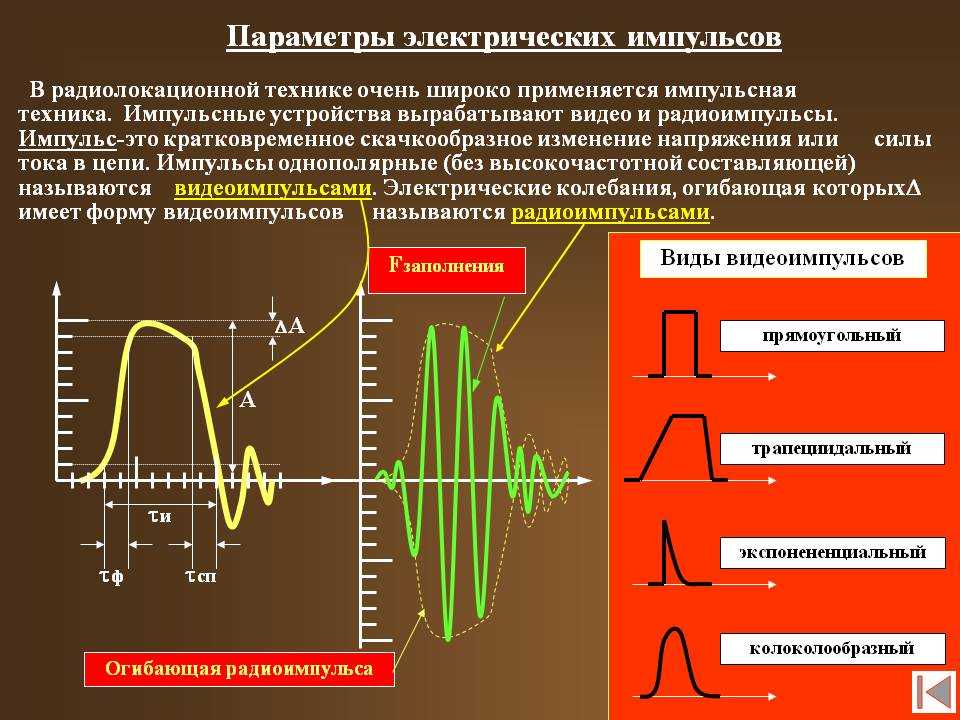 Но мы можем оценивать расстояние куда быстрее и это расстояние в каждый момент времени четко определено.
Но мы можем оценивать расстояние куда быстрее и это расстояние в каждый момент времени четко определено.
Абсолютно то же самое происходит и с другой информацией, мы можем оценивать громкость в любой момент, чувствовать какое давление наши пальцы оказывают на предметы и т.п. Иными словами, практически вся информация, которая может возникать в природе имеет аналоговый вид. Передавать подобную информацию проще всего аналоговыми сигналами, которые являются непрерывными и определены в любой момент времени.
Чтобы понять, как выглядит аналоговый электрический сигнал, можно представить себе график, на котором будет отображена амплитуда по вертикальной оси и время по горизонтальной оси. Если мы, к примеру, замеряем изменение температуры, то на графике появится непрерывная линия, отображающая ее значение в каждый момент времени. Чтобы передать такой сигнал с помощью электрического тока, нам надо сопоставить значение температуры со значением напряжения. Так, например, 35.342 градуса по Цельсию могут быть закодированы как напряжение 3. 5342 В.
5342 В.
Аналоговые сигналы раньше использовались во всех видах связи. Чтобы избежать помех такой сигнал нужно усиливать. Чем выше уровень шума, то есть помех, тем сильнее надо усиливать сигнал, чтобы его можно было принять без искажения. Такой метод обработки сигнала затрачивает много энергии на выделение тепла. При этом усиленный сигнал может сам стать причиной помех для других каналов связи.
Сейчас аналоговые сигналы еще применяются в телевидении и радио, для преобразования входного сигнала в микрофонах. Но, в целом, этот тип сигнала повсеместно вытеснен или вытесняется цифровыми сигналами.
Цифровой сигналЦифровой сигнал представлен последовательностью цифровых значений. Чаще всего сейчас применяются двоичные цифровые сигналы, так как они используются в двоичной электронике и легче кодируются.
В отличие от предыдущего типа сигнала цифровой сигнал имеет два значения «1» и «0». Если мы вспомним наш пример с измерением температуры, то тут сигнал будет сформирован иначе. Если напряжение, которое подается аналоговым сигналом соответствует значению измеряемой температуры, то в цифровом сигнале для каждого значения температуры будет подаваться определенное количество импульсов напряжения. Сам импульс напряжения тут будет равен «1», а отсутствие напряжения – «0». Приемная аппаратура будет декодировать импульсы и восстановит исходные данные.
Если напряжение, которое подается аналоговым сигналом соответствует значению измеряемой температуры, то в цифровом сигнале для каждого значения температуры будет подаваться определенное количество импульсов напряжения. Сам импульс напряжения тут будет равен «1», а отсутствие напряжения – «0». Приемная аппаратура будет декодировать импульсы и восстановит исходные данные.
Представив, как будет выглядеть цифровой сигнал на графике, мы увидим, что переход от нулевого значения к максимальному производится резко. Именно эта особенность позволяет принимающей аппаратуре более четко «видеть» сигнал. Если возникают какие-либо помехи, приемнику проще декодировать сигнал, нежели чем при аналоговой передаче.
Однако цифровой сигнал с очень большим уровнем шума восстановить невозможно, тогда как из аналогового типа при большом искажении еще есть возможность «выудить» информацию. Это связано с эффектом обрыва. Суть эффекта в том, что цифровые сигналы могут передаваться на определенные расстояния, а затем просто обрываются. Этот эффект возникает повсеместно и решается простой регенерацией сигнала. Там, где сигнал обрывается, нужно вставить повторитель или уменьшить длину линии связи. Повторитель не усиливает сигнал, а распознает его изначальный вид и выдает его точную копию и может использоваться сколь угодно в цепи. Такие способы повторения сигнала активно применяются в сетевых технологиях.
Этот эффект возникает повсеместно и решается простой регенерацией сигнала. Там, где сигнал обрывается, нужно вставить повторитель или уменьшить длину линии связи. Повторитель не усиливает сигнал, а распознает его изначальный вид и выдает его точную копию и может использоваться сколь угодно в цепи. Такие способы повторения сигнала активно применяются в сетевых технологиях.
Помимо всего прочего аналоговый и цифровой сигнал различается и возможность кодирования и шифрования информации. Это является одной из причин перехода мобильной связи на «цифру».
Аналоговый и цифровой сигнал и цифро-аналоговое преобразованияСледует еще немного рассказать о том, как аналоговая информация передается по цифровым каналам связи. Вновь прибегнем к примерам. Как уже говорилось звук – это аналоговый сигнал.
Что происходит в мобильных телефонах, которые передают информацию по цифровым каналам
Звук, попадая в микрофон подвергается аналого-цифровому преобразованию (АЦП).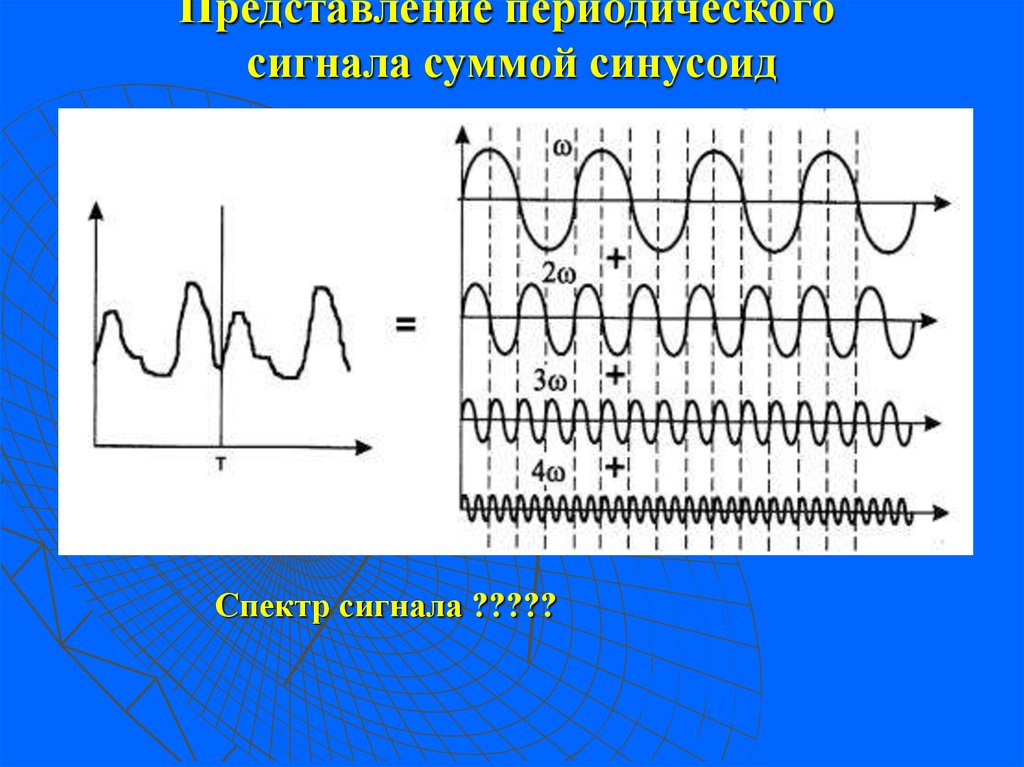 Этот процесс состоит из 3 ступеней. Берутся отдельные значения сигнала через одинаковые отрезки времени, этот процесс называется дискретизация. По теореме Котельникова о пропускной способности каналов, частота взятия этих значений должна быть вдвое выше, чем самая высокая частота сигнала. То есть, если в нашем канале стоит ограничение на частоту в 4 кГц, то частота дискретизации будет составлять 8 кГц.
Этот процесс состоит из 3 ступеней. Берутся отдельные значения сигнала через одинаковые отрезки времени, этот процесс называется дискретизация. По теореме Котельникова о пропускной способности каналов, частота взятия этих значений должна быть вдвое выше, чем самая высокая частота сигнала. То есть, если в нашем канале стоит ограничение на частоту в 4 кГц, то частота дискретизации будет составлять 8 кГц.
Далее все выбранные значения сигнала округляются или, иначе говоря, квантуются. Чем больше уровней при этом будет создано, тем выше будет точность восстановленного сигнала на приемнике. Затем все значения преобразуются в двоичный код, который передается на базовую станцию и затем доходит до другого абонента, являющегося приемником. В телефоне приемника происходит процедура цифро-аналогового преобразования (ЦАП). Это обратная процедура, цель которой на выходе получить сигнал как можно более идентичный исходному. Далее уже аналоговый сигнал выходит в виде звука из динамика телефона.
Каждый день люди сталкиваются с использованием электронных приборов. Без них невозможна современная жизнь. Ведь речь идет о телевизоре, радио, компьютере, телефоне, мультиварке и прочем. Раньше, еще несколько лет назад, никто не задумывался о том, какой сигнал используется в каждом работоспособном приборе. Сейчас же слова «аналоговый», «цифровой», «дискретный» уже давно на слуху. Некоторые виды сигналов из перечисленных являются качественными и надежными.
Без них невозможна современная жизнь. Ведь речь идет о телевизоре, радио, компьютере, телефоне, мультиварке и прочем. Раньше, еще несколько лет назад, никто не задумывался о том, какой сигнал используется в каждом работоспособном приборе. Сейчас же слова «аналоговый», «цифровой», «дискретный» уже давно на слуху. Некоторые виды сигналов из перечисленных являются качественными и надежными.
Цифровая передача стала использоваться намного позже, чем аналоговая. Это связано с тем, что такой сигнал намного проще обслуживать, да и техника на тот момент не была настолько усовершенствована.
С понятием «дискретность» сталкивается каждый человек постоянно. Если переводить это слово с латинского языка, то означать оно будет «прерывистость». Углубляясь далеко в науку, можно сказать, что дискретный сигнал представляет собой метод передачи информации, который подразумевает изменение во времени среды-переносчика. Последняя принимает любое значение из всех возможных. Сейчас дискретность уходит на второй план, после того, как было принято решение производить системы на чипе. Они являются целостными, а все компоненты тесно взаимодействуют друг с другом. В дискретности же все с точностью наоборот – каждая деталь завершена и связана с другими за счет специальных линий связи.
Они являются целостными, а все компоненты тесно взаимодействуют друг с другом. В дискретности же все с точностью наоборот – каждая деталь завершена и связана с другими за счет специальных линий связи.
Сигнал
Сигнал представляет собой специальный код, который передается в пространство одной или несколькими системами. Эта формулировка является общей.
В сфере информации и связи сигналом назван специальный носитель каких-либо данных, который используется для передачи сообщений. Он может быть создан, но не принят, последнее условие не обязательно. Если же сигнал является сообщением, то его «ловля» считается необходимой.
Описываемый код передачи данных задается математической функцией. Она характеризует все возможные изменения параметров. В радиотехнической теории эта модель считается базовой. В ней же аналогом сигнала был назван шум. Он представляет собой функцию времени, которая свободно взаимодействует с переданным кодом и искажает его.
В статье охарактеризованы виды сигналов: дискретный, аналоговый и цифровой. Также коротко дана основная теория по описываемой теме.
Также коротко дана основная теория по описываемой теме.
Виды сигналов
Существует несколько типов классификации имеющихся сигналов. Рассмотрим, какие бывают виды.
- По физической среде носителя данных разделяют электрический сигнал, оптический, акустический и электромагнитный. Имеется еще несколько видов, однако они малоизвестны.
- По способу задания сигналы делятся на регулярные и нерегулярные. Первые представляют собой детерминированные методы передачи данных, которые задаются аналитической функцией. Случайные же формулируются за счет теории вероятности, а также они принимают любые значения в различные промежутки времени.
- В зависимости от функций, которые описывают все параметры сигнала, методы передачи данных могут быть аналоговыми, дискретными, цифровыми (способ, который является квантованным по уровню). Они используются для обеспечения работы многих электрических приборов.
Теперь читателю известны все виды передачи сигналов. Разобраться в них не составит труда любому человеку, главное — немного подумать и вспомнить школьный курс физики.
Разобраться в них не составит труда любому человеку, главное — немного подумать и вспомнить школьный курс физики.
Для чего обрабатывается сигнал?
Сигнал обрабатывается с целью передачи и получения информации, которая в нем зашифрована. Как только она будет извлечена, ее можно использовать различными способами. В отдельных ситуациях ее переформатируют.
Существует и другая причина обработки всех сигналов. Она заключается в небольшом сжатии частот (чтобы не повредить информацию). После этого ее форматируют и передают на медленных скоростях.
В аналоговом и цифровом сигналах используются особенные методы. В частности, фильтрация, свертка, корреляция. Они необходимы для восстановления сигнала, если он поврежден или имеет шум.
Создание и формирование
Зачастую для формирования сигналов необходим аналого-цифровой (АЦП) и цифро-аналоговый (ЦАП) преобразователи. Чаще всего они оба используются лишь в ситуации с применением DSP-технологий. В остальных случаях подойдет только использование ЦАП.
При создании физических аналоговых кодов с дальнейшим применением цифровых методов полагаются на полученную информацию, которая передается со специальных приборов.
Динамический диапазон
Диапазон сигнала вычисляется разностью большего и меньшего уровня громкости, которые выражены в децибелах. Он полностью зависит от произведения и особенностей исполнения. Речь идет как о музыкальных треках, так и об обычных диалогах между людьми. Если брать, например, диктора, который читает новости, то его динамический диапазон колеблется в районе 25-30 дБ. А во время чтения какого-либо произведения он может вырастать до 50 дБ.
Аналоговый сигнал
Аналоговый сигнал является непрерывным во времени способом передачи данных. Недостатком его можно назвать присутствие шума, который иногда приводит к полной потере информации. Очень часто возникают такие ситуации, что невозможно определить, где в коде важные данные, а где обычные искажения.
Именно из-за этого цифровая обработка сигналов приобрела большую популярность и постепенно вытесняет аналоговую.
Цифровой сигнал
Цифровой сигнал является особым потоком данных, он описывается за счет дискретных функций. Его амплитуда может принять определенное значение из уже заданных. Если аналоговый сигнал способен поступать с огромным количеством шумов, то цифровой отфильтровывает большую часть полученных помех.
Помимо этого, такой вид передачи данных переносит информацию без лишней смысловой нагрузки. Через один физический канал может быть отправлено сразу несколько кодов.
Виды цифрового сигнала не существуют, так как он выделяется как отдельный и самостоятельный метод передачи данных. Он представляет собой двоичный поток. В наше время такой сигнал считается самым популярным. Это связано с простотой использования.
Применение цифрового сигнала
Чем же отличается цифровой электрический сигнал от других? Тем, что он способен совершать в ретрансляторе полную регенерацию. Когда в оборудование связи поступает сигнал, имеющий малейшие помехи, он сразу же меняет свою форму на цифровую.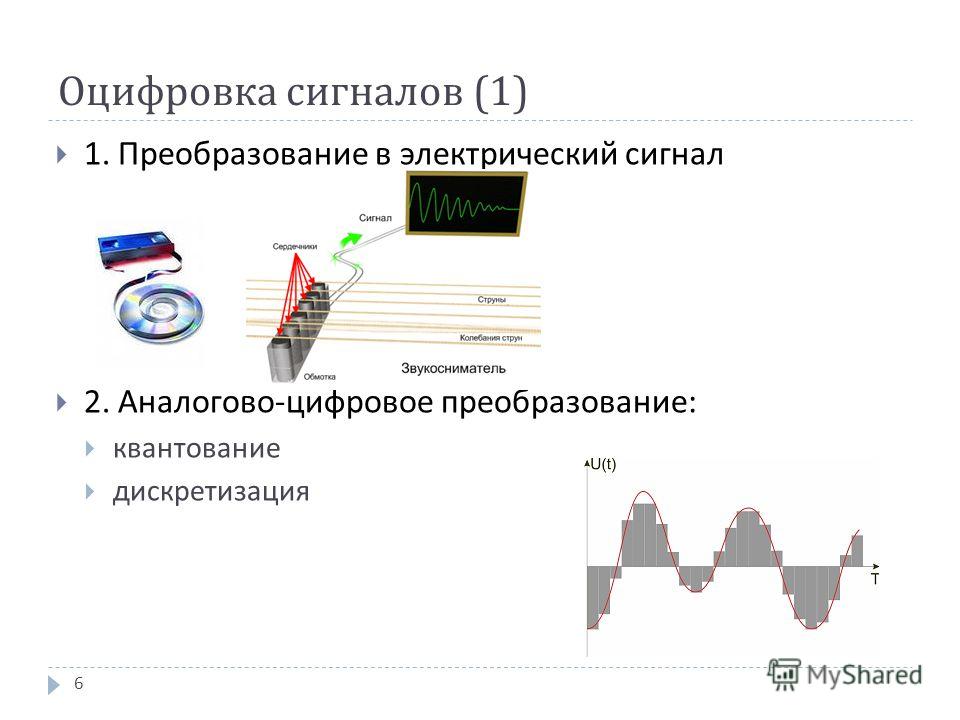 Это позволяет, например, телевышке снова сформировать сигнал, но уже без шумового эффекта.
Это позволяет, например, телевышке снова сформировать сигнал, но уже без шумового эффекта.
В том случае, если код поступает уже с большими искажениями, то, к сожалению, восстановлению он не подлежит. Если брать в сравнении аналоговую связь, то в аналогичной ситуации ретранслятор может извлечь часть данных, затрачивая много энергии.
Обсуждая сотовую связь разных форматов, при сильном искажении на цифровой линии разговаривать практически невозможно, так как не слышны слова или целые фразы. Аналоговая связь в таком случае более действенна, ведь можно продолжать вести диалог.
Именно из-за подобных неполадок цифровой сигнал ретрансляторы формируют очень часто для того, чтобы сократить разрыв линии связи.
Дискретный сигнал
Сейчас каждый человек пользуется мобильным телефоном или какой-то «звонилкой» на своем компьютере. Одна из задач приборов или программного обеспечения – это передача сигнала, в данном случае голосового потока. Для переноса непрерывной волны необходим канал, который имел бы пропускную способность высшего уровня. Именно поэтому было предпринято решение использовать дискретный сигнал. Он создает не саму волну, а ее цифровой вид. Почему же? Потому что передача идет от техники (например, телефона или компьютера). В чем плюсы такого вида переноса информации? С его помощью уменьшается общее количество передаваемых данных, а также легче организуется пакетная отправка.
Именно поэтому было предпринято решение использовать дискретный сигнал. Он создает не саму волну, а ее цифровой вид. Почему же? Потому что передача идет от техники (например, телефона или компьютера). В чем плюсы такого вида переноса информации? С его помощью уменьшается общее количество передаваемых данных, а также легче организуется пакетная отправка.
Понятие «дискретизация» уже давно стабильно используется в работе вычислительной техники. Благодаря такому сигналу передается не непрерывная информация, которая полностью закодирована специальными символами и буквами, а данные, собранные в особенные блоки. Они являются отдельными и законченными частицами. Такой метод кодировки уже давно отодвинулся на второй план, однако не исчез полностью. С помощью него можно легко передавать небольшие куски информации.
Сравнение цифрового и аналогового сигналов
Покупая технику, вряд ли кто-то думает о том, какие виды сигналов использованы в том или другом приборе, а об их среде и природе уж тем более.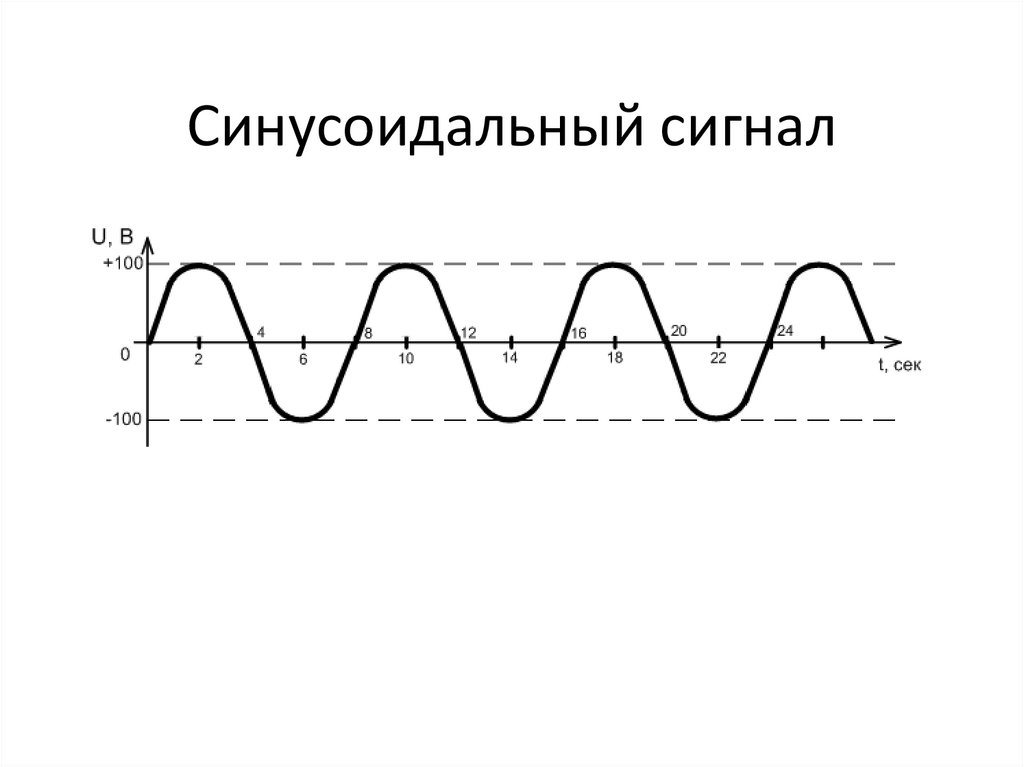 Но иногда все же приходится разбираться с понятиями.
Но иногда все же приходится разбираться с понятиями.
Уже давно стало ясно, что аналоговые технологии теряют спрос, ведь их использование нерационально. Взамен приходит цифровая связь. Нужно понимать, о чем идет речь и от чего отказывается человечество.
Если говорить коротко, то аналоговый сигнал – способ передачи информации, который подразумевает описание данных непрерывными функциями времени. По сути, говоря конкретно, амплитуда колебаний может быть равна любому значению, находящемуся в определенных границах.
Цифровая обработка сигналов описывается дискретными функциями времени. Иначе говоря, амплитуда колебаний этого метода равна строго заданным значениям.
Переходя от теории к практике, надо сказать о том, что аналоговому сигналу характерны помехи. С цифровым же таких проблем нет, потому что он успешно их «сглаживает». За счет новых технологий такой метод передачи данных способен своими силами без вмешательства ученого восстановить всю исходную информацию.
Говоря о телевидении, можно уже с уверенностью сказать: аналоговая передача давно изжила себя.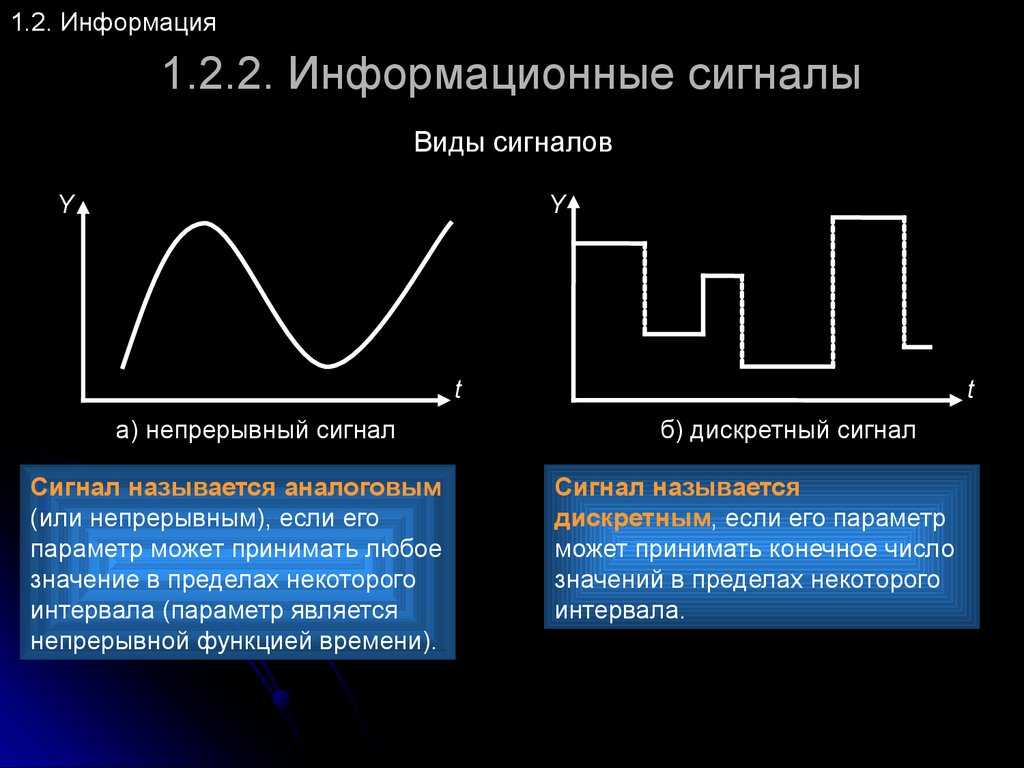 Большинство потребителей переходят на цифровой сигнал. Минус последнего заключается в том, что если аналоговую передачу способен принимать любой прибор, то более современный способ – только специальная техника. Хоть и спрос на устаревший метод уже давно упал, все же такие виды сигналов до сих пор не способны полностью уйти из повседневной жизни.
Большинство потребителей переходят на цифровой сигнал. Минус последнего заключается в том, что если аналоговую передачу способен принимать любой прибор, то более современный способ – только специальная техника. Хоть и спрос на устаревший метод уже давно упал, все же такие виды сигналов до сих пор не способны полностью уйти из повседневной жизни.
Электрические сигналы мозга доступны для понимания. Глава из книги «Мозговой трест» – аналитический портал ПОЛИТ.РУ
Издательство «Синдбад» представляет книгу «Мозговой трест. 39 ведущих нейробиологов — о том, что мы знаем и чего не знаем о мозге» (под редакцией Дэвида Линдена, перевод Юрия Гольдберга).
Профессор Дэвид Линден собрал ответы тридцати девяти ведущих нейробиологов на вопрос «Что бы вы больше всего хотели рассказать людям о работе мозга?». Так родился этот сборник научно-популярных эссе, расширяющий представление о человеческом мозге и его возможностях. В нем специалисты по человеческому поведению, молекулярной генетике, эволюционной биологии и сравнительной анатомии освещают самые разные темы.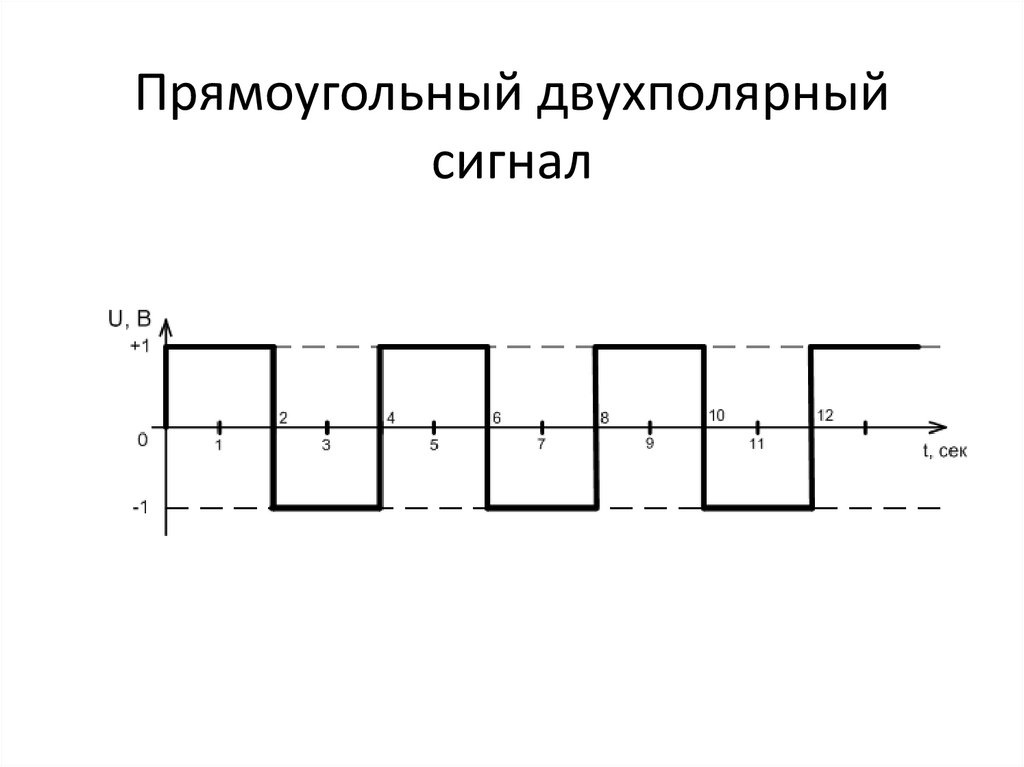 Почему время в нашем восприятии то летит незаметно, то тянется бесконечно долго? Почему, управляя автомобилем, мы ощущаем его частью своего тела? Почему дети осваивают многие навыки быстрее взрослых? Что творится в голове у подростка? Какой механизм отвечает за нашу интуицию? Способны ли мы читать чужие мысли? Как биологические факторы влияют на сексуальную ориентацию? Как меняется мозг под воздействием наркотиков? Как помочь мозгу восстановиться после инсульта? Наконец, возможно ли когда-нибудь создать искусственный мозг, подобный человеческому? Авторы описывают самые удивительные особенности мозга, честно объясняя, что известно, а что пока неизвестно ученым о работе нервной системы.
Почему время в нашем восприятии то летит незаметно, то тянется бесконечно долго? Почему, управляя автомобилем, мы ощущаем его частью своего тела? Почему дети осваивают многие навыки быстрее взрослых? Что творится в голове у подростка? Какой механизм отвечает за нашу интуицию? Способны ли мы читать чужие мысли? Как биологические факторы влияют на сексуальную ориентацию? Как меняется мозг под воздействием наркотиков? Как помочь мозгу восстановиться после инсульта? Наконец, возможно ли когда-нибудь создать искусственный мозг, подобный человеческому? Авторы описывают самые удивительные особенности мозга, честно объясняя, что известно, а что пока неизвестно ученым о работе нервной системы.
Предлагаем прочитать главу, написанную Дэвидом Фостером из Калифорнийского университета в Беркли.
Электрические сигналы мозга доступны для понимания
Дэвид Фостер
Это история о том, как ученые использовали электроды для регистрации активности нейронов мозга, чтобы узнать, как они реагируют не только на визуальные образы, запахи и звуки, но и на память, эмоции, удовольствие и боль.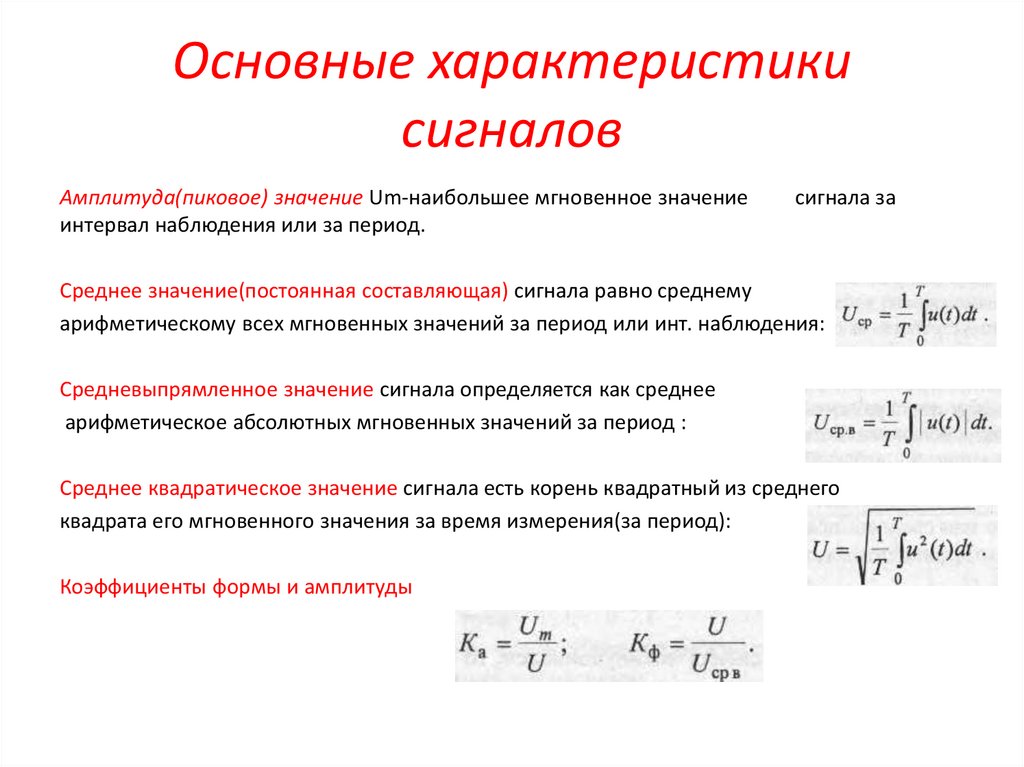 Нейроны — особенные клетки: они связываются с десятками тысяч других нейронов и образуют разветвленные сети, отвечающие за сложные вычисления, которые лежат в основе всех наших действий. Эта история охватывает столетие удивительных открытий, череду периодов оптимизма и пессимизма, а также открывает новый поразительный факт: электрические сигналы, вырабатываемые нейронами, можно понять.
Нейроны — особенные клетки: они связываются с десятками тысяч других нейронов и образуют разветвленные сети, отвечающие за сложные вычисления, которые лежат в основе всех наших действий. Эта история охватывает столетие удивительных открытий, череду периодов оптимизма и пессимизма, а также открывает новый поразительный факт: электрические сигналы, вырабатываемые нейронами, можно понять.
Самые ранние открытия показали, что каждый нейрон передает электрический импульс, или спайк, другим нейронам, с которыми он контактирует. Считается, что эта передача осуществляется по принципу «всё или ничего». Не бывает ни «громких», ни «тихих» спайков: спайк — это всегда просто спайк. Мозг использует эти импульсы для отправки своеобразного кода, в котором объекты и события внешнего мира представляются на удивление простым и понятным образом. Например, сила стимуляции периферийного нерва, реагирующего на прикосновение к пальцу, передается в виде некоторого числа спайков, поступающих от нерва за определенный период времени. В сетчатке глаза лягушки были найдены нейроны, которые возбуждаются только в том случае, когда в определенной части поля зрения появляется маленький черный диск; активация этих «детекторов жуков» вызывает хищническую поведенческую реакцию лягушки в направлении диска. В 1950-е и 1960-е годы, в золотой век записи сигналов отдельных нейронов, будущие лауреаты Нобелевской премии Дэвид Хьюбел и Торстен Визель обнаружили, что нейроны в зрительной коре головного мозга реагируют на такие элементы визуальной картины, как резкие границы. Дальнейшие исследования позволили выявить в более глубоких отделах мозга нейроны, настроенные на более сложные элементы. Кульминацией этих открытий стало обнаружение нейронов, избирательно реагирующих на руки и лица.
В сетчатке глаза лягушки были найдены нейроны, которые возбуждаются только в том случае, когда в определенной части поля зрения появляется маленький черный диск; активация этих «детекторов жуков» вызывает хищническую поведенческую реакцию лягушки в направлении диска. В 1950-е и 1960-е годы, в золотой век записи сигналов отдельных нейронов, будущие лауреаты Нобелевской премии Дэвид Хьюбел и Торстен Визель обнаружили, что нейроны в зрительной коре головного мозга реагируют на такие элементы визуальной картины, как резкие границы. Дальнейшие исследования позволили выявить в более глубоких отделах мозга нейроны, настроенные на более сложные элементы. Кульминацией этих открытий стало обнаружение нейронов, избирательно реагирующих на руки и лица.
Триумфальную атмосферу той эпохи довольно точно передает статья физиолога Хораса Барлоу, который сформулировал однонейронную теорию восприятия. Барлоу утверждал, что восприятие — это активность одного нейрона и что всю психику человека можно объяснить с точки зрения такой активности. Однако нейронов, настроенных на какую-то конкретную зрительную составляющую, находили все меньше и меньше, что поставило под сомнение эту теорию. Дэвид Марр был блестящим ученым, который первым начал разрабатывать математические модели возможной работы нейронных сетей. (К сожалению, он умер от лейкемии в возрасте 35 лет.) Марр написал знаменитую книгу «Зрение», опубликованную в 1982 году, уже после его смерти. В вводной главе он поставил под сомнение существующие взгляды на связь нервной активности и психических процессов. Вот как он рассуждает о нейроне с избирательной активностью, который реагирует только на присутствие бабушки:
Однако нейронов, настроенных на какую-то конкретную зрительную составляющую, находили все меньше и меньше, что поставило под сомнение эту теорию. Дэвид Марр был блестящим ученым, который первым начал разрабатывать математические модели возможной работы нейронных сетей. (К сожалению, он умер от лейкемии в возрасте 35 лет.) Марр написал знаменитую книгу «Зрение», опубликованную в 1982 году, уже после его смерти. В вводной главе он поставил под сомнение существующие взгляды на связь нервной активности и психических процессов. Вот как он рассуждает о нейроне с избирательной активностью, который реагирует только на присутствие бабушки:
Допустим, например, что кому-то и правда удалось обнаружить мифическую «нервную клетку для бабушки». Даст ли нам такое открытие что-нибудь действительно важное? Мы будем знать, что такая клетка существует (гроссовские детекторы руки значат для нас практически именно это), но не будем знать, зачем или хотя бы каким образом подобный феномен может быть синтезирован из выходных сигналов уже известных нервных клеток.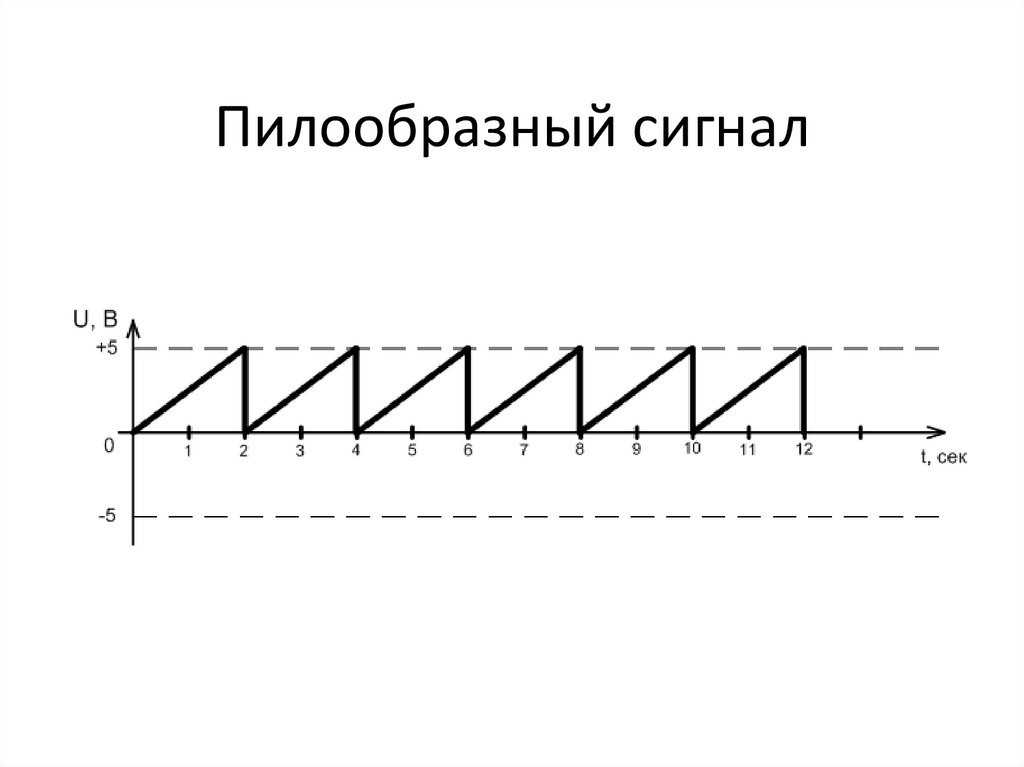 .. Если бы нам действительно стали известны ответы на эти вопросы, можно было бы, например, использовать их в программе вычислительной машины. Обнаружение детектора руки, однако, явно не дает возможности написать программу, реализующую такой детектор.
.. Если бы нам действительно стали известны ответы на эти вопросы, можно было бы, например, использовать их в программе вычислительной машины. Обнаружение детектора руки, однако, явно не дает возможности написать программу, реализующую такой детектор.
Отбросив чисто эмпирический подход, Марр предложил нечто совершенно новое: изучить, как должна быть устроена зрительная система с инженерной точки зрения. Для этого необходимо использовать знания из области искусственного интеллекта (ИИ). Эти знания позволят разработать эффективные компьютерные программы, и тогда — и только тогда — появится возможность интерпретировать особенности нейронных реакций.
Парадоксально, что Марр призывал использовать ИИ в нейробиологии как раз в тот момент, когда энтузиазм в отношении ИИ пошел на спад. Грандиозная задача разработать программы для распознавания речи и образов оказалась слишком трудной. Выяснилось, что эти действия, с которыми мы справляемся без труда, очень сложно описать в виде последовательных шагов программы. Марр хотел, чтобы нейробиология остановилась и подождала, пока ИИ не предложит решение, но в развитии ИИ произошел неожиданный поворот. Был разработан новый тип вычислений на основе сетей искусственных нейронов, которые не программировались на решение тех или иных задач, а постепенно учились на примерах. Эта сфера также пережила волны оптимизма и пессимизма, но к настоящему времени глубокое обучение позволило ИИ почти догнать человека в решении задач, которые раньше казались невыполнимыми: распознавание лиц и управление автомобилем.
Марр хотел, чтобы нейробиология остановилась и подождала, пока ИИ не предложит решение, но в развитии ИИ произошел неожиданный поворот. Был разработан новый тип вычислений на основе сетей искусственных нейронов, которые не программировались на решение тех или иных задач, а постепенно учились на примерах. Эта сфера также пережила волны оптимизма и пессимизма, но к настоящему времени глубокое обучение позволило ИИ почти догнать человека в решении задач, которые раньше казались невыполнимыми: распознавание лиц и управление автомобилем.
Тем временем нейрофизиологи продолжали делать важные открытия, связанные с реакцией нейронов, особенно в тех областях мозга, которые не ассоциировались с первичной обработкой сенсорной информации. Выяснилось, что нейроны в глубине медиальной височной доли и в гиппокампе участвуют в формировании памяти, получая информацию об окружающем мире, предварительно обработанную другими отделами мозга, куда она поступает от глаз и других органов чувств; аналогичным образом исходящие сигналы гиппокампа проходят через разные области, прежде чем вызвать какие-либо действия. Гиппокамп по-прежнему представлял собой загадочную область мозга, и мало кто ожидал, что наблюдения выявят значимую и интуитивно понятную корреляцию в поведении этих нейронов. Однако сочетание новых экспериментальных методов и философского подхода позволило Джону О’Кифу и его коллегам в начале 1970-х найти эту корреляцию. Позволив крысам свободно исследовать окружающую среду, О’Киф обнаружил, что нейроны их гиппокампа генерируют импульсы только в определенных областях пространства, разных для каждого нейрона. Основываясь на идее Канта об априорном знании, О’Киф и психолог Линн Нейдел предположили, что эти нейроны предоставляют остальному мозгу когнитивную карту пространства, и эта идея вдохновила целое поколение исследователей (как сторонников, так и противников гипотезы).
Гиппокамп по-прежнему представлял собой загадочную область мозга, и мало кто ожидал, что наблюдения выявят значимую и интуитивно понятную корреляцию в поведении этих нейронов. Однако сочетание новых экспериментальных методов и философского подхода позволило Джону О’Кифу и его коллегам в начале 1970-х найти эту корреляцию. Позволив крысам свободно исследовать окружающую среду, О’Киф обнаружил, что нейроны их гиппокампа генерируют импульсы только в определенных областях пространства, разных для каждого нейрона. Основываясь на идее Канта об априорном знании, О’Киф и психолог Линн Нейдел предположили, что эти нейроны предоставляют остальному мозгу когнитивную карту пространства, и эта идея вдохновила целое поколение исследователей (как сторонников, так и противников гипотезы).
Внутри узкого круга нейрофизиологов, изучавших гиппокамп, разгорелись споры о том, в какой степени реакция нейронов места в гиппокампе отображает окружающее пространство. Эта реакция не выглядела достаточно регулярной, чтобы доказывать геометрическое отображение, которое хотели бы приписать им некоторые исследователи.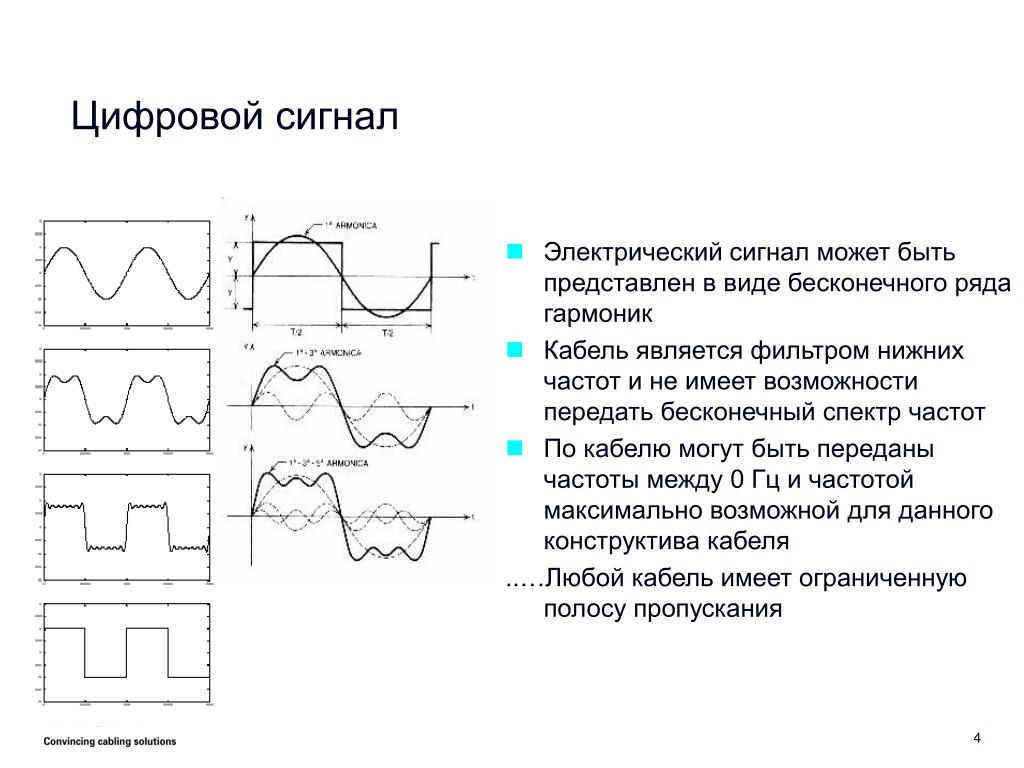 В итоге в нескольких областях, связанных с гиппокампом, были найдены нейроны, обладающие именно такими «геометрическими» свойствами. Сначала были обнаружены нейроны, которые действовали как внутренний компас и реагировали на направление, в котором поворачивалось животное. Затем в 2005 году Эдвард и Мэй-Бритт Мозер с коллегами сообщили, что нейроны энторинальной коры, в которую поступают исходящие сигналы гиппокампа, реагируют на шестиугольную сетку в пространстве — то есть на определенный узор, для которого характерна периодичность в двух измерениях. Удивительно, но эти нейроны решетки не связаны с поведенческими траекториями животных; скорее, они отражают внутреннюю структуру, которая соответствует окружающему пространству, нечто вроде схемы на бумаге. В 2014 году О’Киф и супруги Мозер за свои открытия были удостоены Нобелевской премии. С тех пор в тех же отделах мозга, а также в других, связанных с ними, были найдены другие нейроны, реагирующие на положение в пространстве.
В итоге в нескольких областях, связанных с гиппокампом, были найдены нейроны, обладающие именно такими «геометрическими» свойствами. Сначала были обнаружены нейроны, которые действовали как внутренний компас и реагировали на направление, в котором поворачивалось животное. Затем в 2005 году Эдвард и Мэй-Бритт Мозер с коллегами сообщили, что нейроны энторинальной коры, в которую поступают исходящие сигналы гиппокампа, реагируют на шестиугольную сетку в пространстве — то есть на определенный узор, для которого характерна периодичность в двух измерениях. Удивительно, но эти нейроны решетки не связаны с поведенческими траекториями животных; скорее, они отражают внутреннюю структуру, которая соответствует окружающему пространству, нечто вроде схемы на бумаге. В 2014 году О’Киф и супруги Мозер за свои открытия были удостоены Нобелевской премии. С тех пор в тех же отделах мозга, а также в других, связанных с ними, были найдены другие нейроны, реагирующие на положение в пространстве.
Несмотря на успехи нейрофизиологии в изучении того, как отдельные клетки отображают внешний мир, многие вопросы по-прежнему оставались без ответов.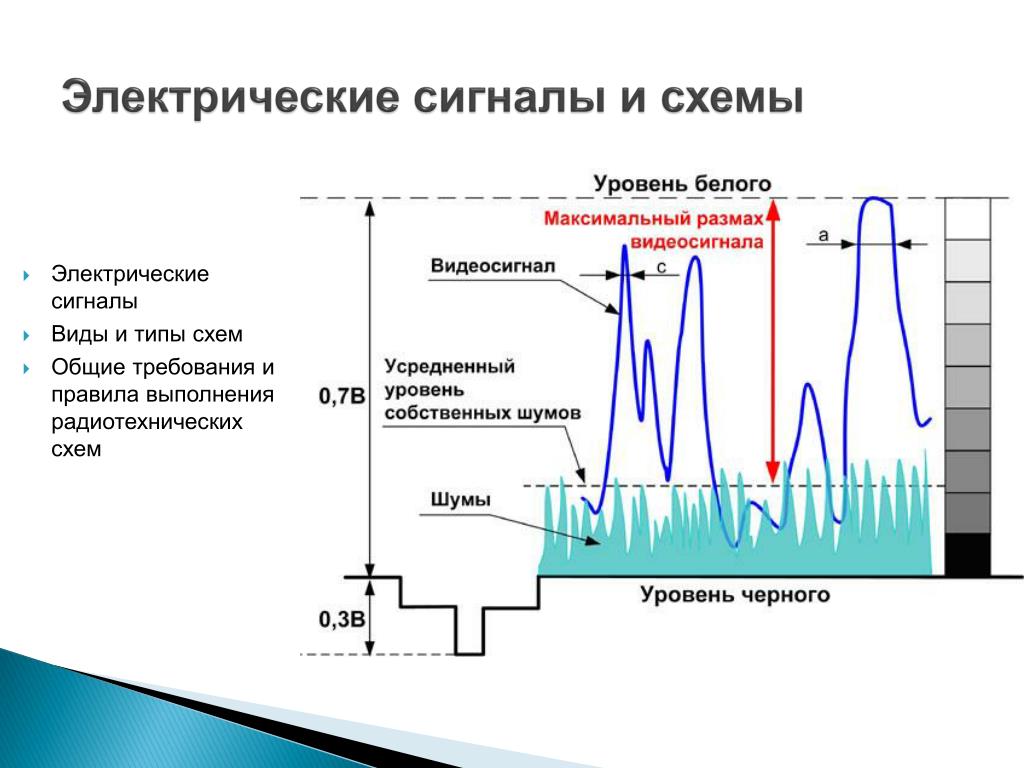 Неужели наши мысли и чувства — всего лишь реакция на стимулы, как и реакции отдельных нейронов? Одно из критических замечаний Марра, которое сохраняет актуальность и по сей день, заключается в том, что знать отображение, осуществляемое одним нейроном, — не значит понимать процесс. Например, мы знаем функцию нейронов места в гиппокампе: они хранят карту окружающего мира; но как именно эта карта используется? Читая карту, вы направляете взгляд назад и вперед, чтобы понять, откуда вы пришли и куда направляетесь. Одно лишь знание о нейронах места ничего не говорит нам о подобных процессах. Как ни странно, ответ могли дать технические достижения в нейрофизиологии. В конце концов, теория одного нейрона, предложенная Барлоу, потребовала сосредоточиться на изучении активности отдельных нейронов, поскольку из-за ограниченных возможностей экспериментальных методов это были единственные доступные данные. Примерно с начала 1990-х стала возможна одновременная регистрация активности большого количества нейронов.
Неужели наши мысли и чувства — всего лишь реакция на стимулы, как и реакции отдельных нейронов? Одно из критических замечаний Марра, которое сохраняет актуальность и по сей день, заключается в том, что знать отображение, осуществляемое одним нейроном, — не значит понимать процесс. Например, мы знаем функцию нейронов места в гиппокампе: они хранят карту окружающего мира; но как именно эта карта используется? Читая карту, вы направляете взгляд назад и вперед, чтобы понять, откуда вы пришли и куда направляетесь. Одно лишь знание о нейронах места ничего не говорит нам о подобных процессах. Как ни странно, ответ могли дать технические достижения в нейрофизиологии. В конце концов, теория одного нейрона, предложенная Барлоу, потребовала сосредоточиться на изучении активности отдельных нейронов, поскольку из-за ограниченных возможностей экспериментальных методов это были единственные доступные данные. Примерно с начала 1990-х стала возможна одновременная регистрация активности большого количества нейронов. Но вот о чем, вероятно, не упоминалось раньше: воображение ученых уводило их совсем не в том направлении, на которое в итоге указали данные.
Но вот о чем, вероятно, не упоминалось раньше: воображение ученых уводило их совсем не в том направлении, на которое в итоге указали данные.
Многие ученые ожидали, что полученные методами параллельной записи данные о реакции популяций нейронов на стимулы будут информативнее данных от одиночных нейронов. Например, предполагалось, что зрительные нейроны, соответствующие границам объекта, будут возбуждаться синхронно, что позволит отличить их от нейронов, соответствующих границам другого объекта. Еще одно предположение заключалось в том, что популяция нейронов может отображать распределение некоторого числа оценок, а не единичную оценку. А вот что оказалось неожиданным: координируя свою активность, нейроны могут действовать независимо от стимула. Эта особенность впервые была замечена у нейронов места в гиппокампе: те из них, которые возбуждались одновременно при том или ином поведении, поскольку отвечали за те же поля места, продолжали одновременно возбуждаться и позже, во сне.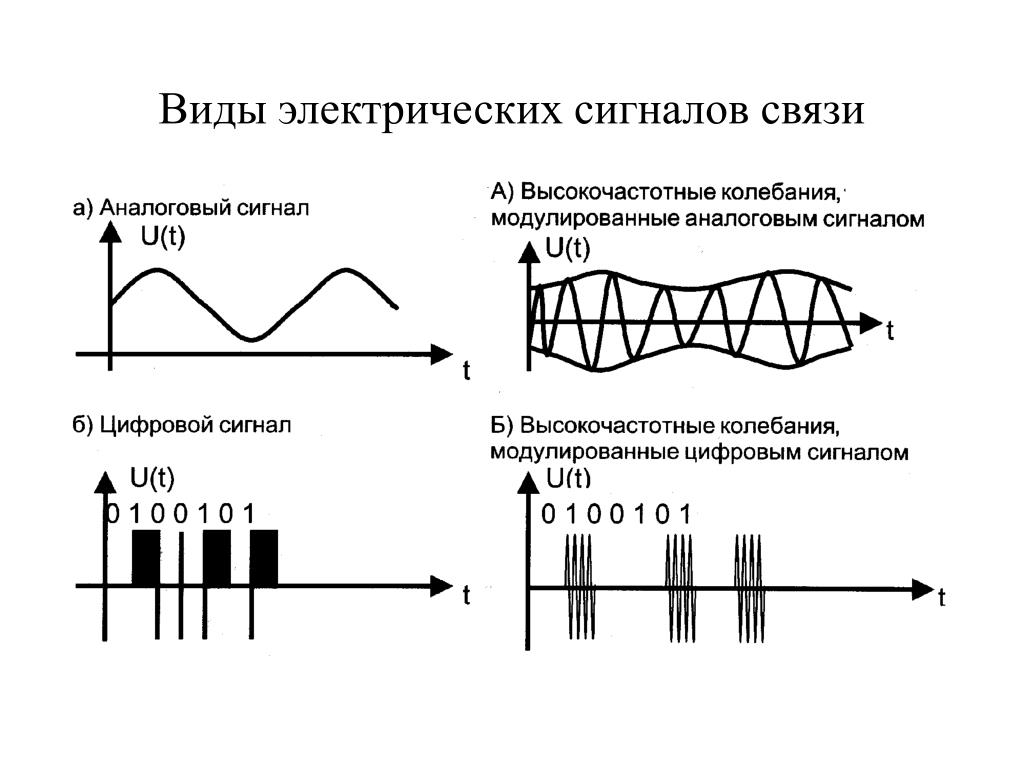 Но это еще не всё. Одновременно записав сигналы от достаточно большого числа клеток, ученые обнаружили, что последовательность активации нейронов места в гиппокампе соответствовала перемещению в окружающей среде — то есть во сне животное как бы повторяло пройденный маршрут, но в ускоренном режиме. Эти последовательности наблюдались не только во сне, но и, что очень важно, когда при решении пространственных задач животные ненадолго останавливались. Создавалось впечатление, что внезапно включалась хранящаяся в гиппокампе карта: луч фонаря (мысленный, без каких бы то ни было движений) сканировал окружающее пространство в поисках возможных поведенческих траекторий — так же, как наши глаза сканируют карту. Теперь мы знаем, что эти последовательности отражают приобретенную в результате обучения информацию о структуре окружающего пространства.
Но это еще не всё. Одновременно записав сигналы от достаточно большого числа клеток, ученые обнаружили, что последовательность активации нейронов места в гиппокампе соответствовала перемещению в окружающей среде — то есть во сне животное как бы повторяло пройденный маршрут, но в ускоренном режиме. Эти последовательности наблюдались не только во сне, но и, что очень важно, когда при решении пространственных задач животные ненадолго останавливались. Создавалось впечатление, что внезапно включалась хранящаяся в гиппокампе карта: луч фонаря (мысленный, без каких бы то ни было движений) сканировал окружающее пространство в поисках возможных поведенческих траекторий — так же, как наши глаза сканируют карту. Теперь мы знаем, что эти последовательности отражают приобретенную в результате обучения информацию о структуре окружающего пространства.
Удивительный факт: в этих последовательностях могут соединяться фрагменты действий, которые никогда не наблюдались вместе, создавая нечто вроде мысленных «коротких путей».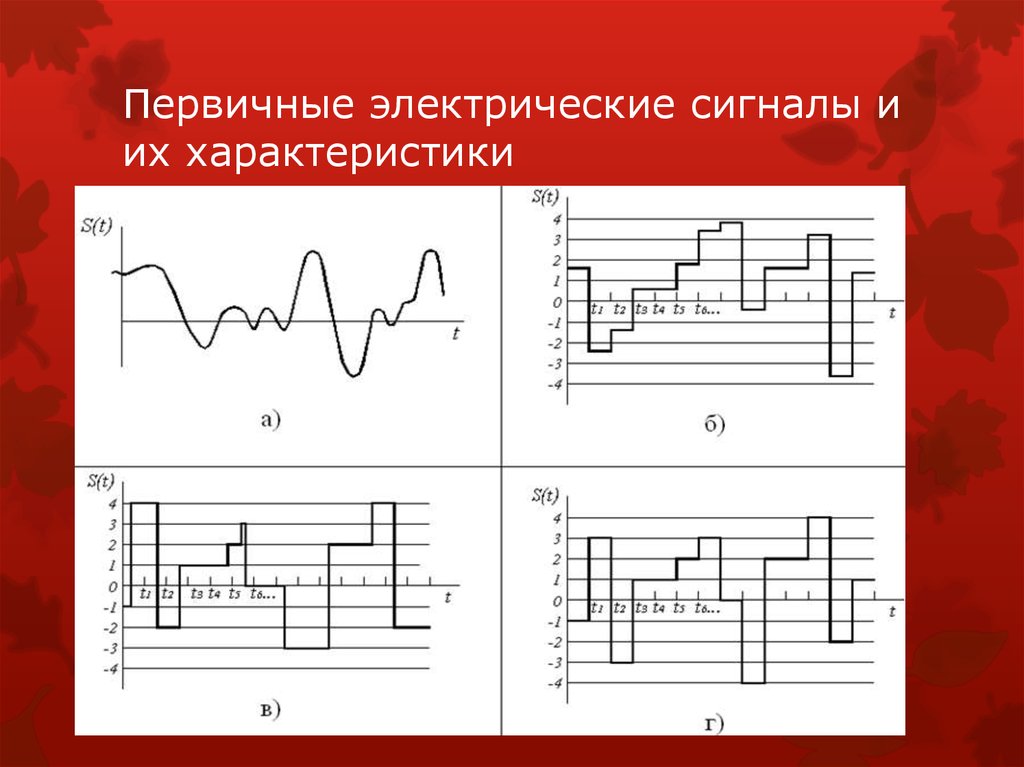 Теперь нам также известно, что последовательности возникают не случайно, а направляются текущим местоположением цели в каждый момент, когда животное намеревается выполнить движение, словно отражая активный процесс выбора пути. Даже классическая активность, записанная во время движения, организована в короткие «упреждающие» последовательности, и этот процесс становится понятным при одновременном исследовании большого числа нейронов места. По сути, большинство спайков, вырабатываемых нейронами гиппокампа, представляют собой части той или иной последовательности. Последовательная организация может иметь базовый характер и присутствовать не только в гиппокампе, но и во всем мозге.
Теперь нам также известно, что последовательности возникают не случайно, а направляются текущим местоположением цели в каждый момент, когда животное намеревается выполнить движение, словно отражая активный процесс выбора пути. Даже классическая активность, записанная во время движения, организована в короткие «упреждающие» последовательности, и этот процесс становится понятным при одновременном исследовании большого числа нейронов места. По сути, большинство спайков, вырабатываемых нейронами гиппокампа, представляют собой части той или иной последовательности. Последовательная организация может иметь базовый характер и присутствовать не только в гиппокампе, но и во всем мозге.
Окончательного вывода из этой истории нет. Мы вступили во второй золотой век нейрофизиологии. Технические достижения, позволяющие наблюдать за большим количеством клеток одновременно и выявлять новые типы клеток, которые раньше были неотличимы друг от друга, произвели революцию в этой области исследований. История не закончена, и то, что сегодня кажется непостижимым, завтра отступит перед результатами экспериментов.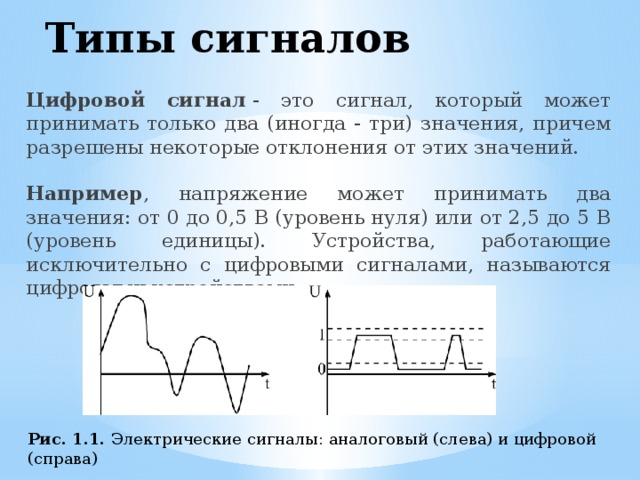 Мозг устроен таким образом, чтобы рисовать последовательности, разворачивающиеся во времени, фантазировать и обдумывать альтернативные планы действий; он ждет, готовясь рассказать о себе историю, которая будет и необычной, и понятной.
Мозг устроен таким образом, чтобы рисовать последовательности, разворачивающиеся во времени, фантазировать и обдумывать альтернативные планы действий; он ждет, готовясь рассказать о себе историю, которая будет и необычной, и понятной.
Интеграция электрических сигналов, возникающих в корнях сосудистых растений
Электрическая природа жизни
Набухание, вызванное давлением, — это проблема, возникшая на ранней стадии эволюции и решенная с появлением мембранных белков, обеспечивающих синхронное перераспределение ионных градиентов через плазматическую мембрану. Вторичным эффектом этого решения является создание падения напряжения внутри мембранного диэлектрика (Finkelstein, 1976; Armstrong, 2015). Активность ионных каналов и транспортеров избирательно регулирует прохождение ионов, вызывая кратковременные локальные изменения мембранного потенциала при включении метаболитов или изменении проницаемости мембраны в ответ на внешний сигнал. С учетом синхронизации клеточных процессов и коммуникации внутри клеточных сообществ электрическая сенсорика и сигнализация развиваются как широко распространенный механизм на разных уровнях биологической организации. От бактериальных биопленок (Strahl, Hamoen, 2010; Masi, Ciszak, 2014; Prindle et al., 2015) до высших растений (Sanderson, 1872; Darwin, 189).7; Бозе, 1907 г .; Pickard, 1973) и животных (Galvani, 1791; Hodgkin, 1937; Cole and Curtis, 1939; Armstrong, 2007) электрическая коммуникация принимает различные формы, различающиеся по своей сложности, от простых градуированных или колебательных изменений мембранного напряжения до передачи электрических сигналов на большие расстояния. наблюдается в возбудимых клетках.
От бактериальных биопленок (Strahl, Hamoen, 2010; Masi, Ciszak, 2014; Prindle et al., 2015) до высших растений (Sanderson, 1872; Darwin, 189).7; Бозе, 1907 г .; Pickard, 1973) и животных (Galvani, 1791; Hodgkin, 1937; Cole and Curtis, 1939; Armstrong, 2007) электрическая коммуникация принимает различные формы, различающиеся по своей сложности, от простых градуированных или колебательных изменений мембранного напряжения до передачи электрических сигналов на большие расстояния. наблюдается в возбудимых клетках.
Электрические сигналы у животных и высших растений
Хорошо известно, что и растения, и животные используют электрические сигналы дальнего действия для передачи информации об окружающей среде всему телу (Armstrong, 2007; Hedrich et al., 2016). В многоклеточных организмах информация должна передаваться от детекторов к эффекторной ткани. В случае животных нервная система играет центральную роль в гомеостазе, выступая в качестве основного интегратора большей части соответствующей физиологической информации.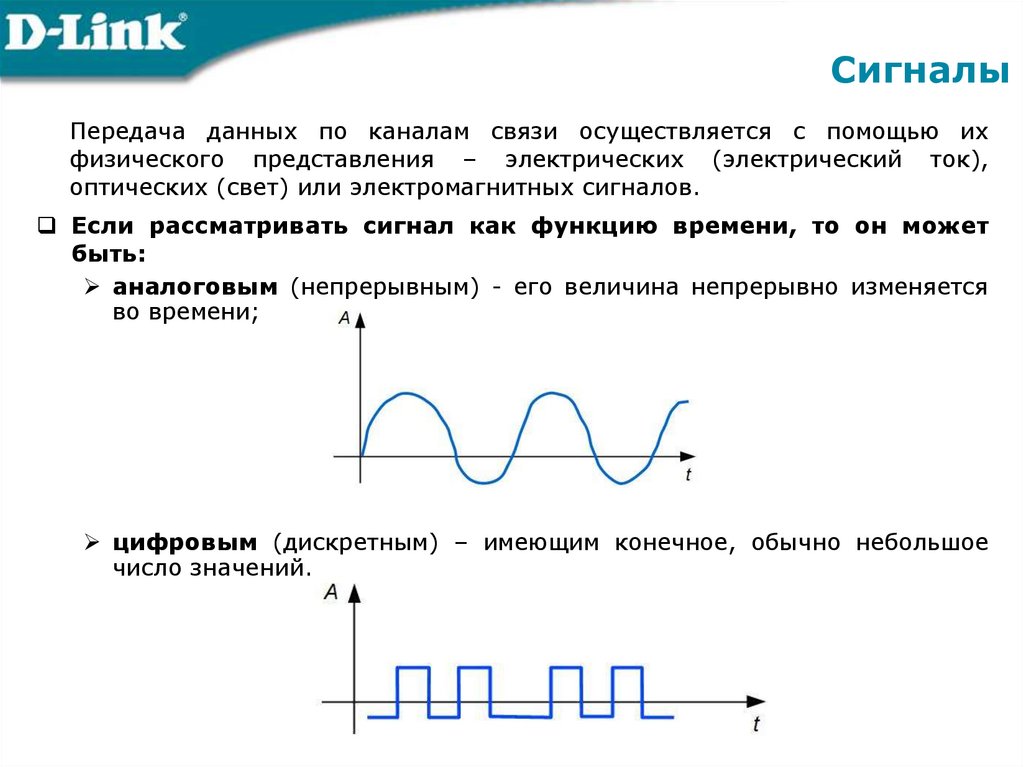 Связь между эпителиальной тканью и возбудимыми клетками определяет то, как животные взаимодействуют с окружающей средой, не только используя преимущества сенсорных модальностей, таких как прикосновение, температура, свет или звук (Frings, 2009).; Julius and Nathans, 2012), но и путем интеграции внутренних процессов, таких как выброс гормонов, физиология кишечника и развитие иммунной системы (Zhang and Zhang, 2009; Bellono et al., 2017; Clemmensen et al., 2017).
Связь между эпителиальной тканью и возбудимыми клетками определяет то, как животные взаимодействуют с окружающей средой, не только используя преимущества сенсорных модальностей, таких как прикосновение, температура, свет или звук (Frings, 2009).; Julius and Nathans, 2012), но и путем интеграции внутренних процессов, таких как выброс гормонов, физиология кишечника и развитие иммунной системы (Zhang and Zhang, 2009; Bellono et al., 2017; Clemmensen et al., 2017).
Молекулярные детекторы, обнаруженные в сенсорном эпителии, активируются сигналами окружающей среды, вызывая (прямо или косвенно) открытие проводимости ионного канала, которое изменяет локальный трансмембранный потенциал (Martinac, 2008). В невозбудимых клетках, таких как эпителиальные клетки кишечника или легких, электрогенный транспорт управляет поглощением питательных веществ, контролирует рН и модулирует секрецию воды (Boyd, 2008; Beumer and Clevers, 2017; Clemmensen et al., 2017). В случае эпителия отсутствие подходящих потенциалзависимых каналов (т.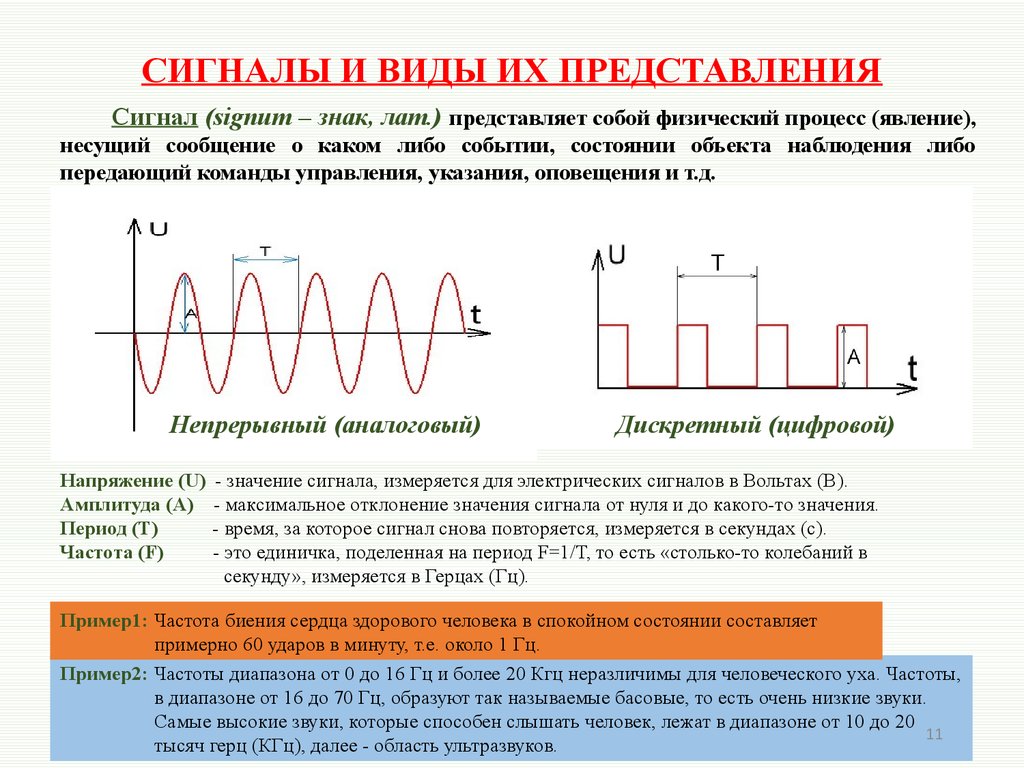 е. Ca v , Na v ) препятствует распространению начальной деполяризации на большие расстояния. В возбудимых клетках деполяризация обеспечивает необходимую энергию для индукции открытия потенциалзависимых каналов (т. е. Ca v с, Na v с и K v с) (Bezanilla, 2008; Catterall et al., 2017). Скорость распространения, форма распространяемого потенциала и частота электрического сообщения определяются свойствами кабеля ячейки, которые определяются как геометрией каждого конкретного типа ячейки, так и набором доступных ионных каналов. У животных этот тип связи распространяется и на многоклеточный организм, когда выделившееся из данной клетки вещество оказывает влияние на постсинаптическую клетку (Gerber, Südhof, 2002; Jackson, 2006; Catterall, Few, 2008). Как описано первоначально в Aplysia Э. Канделя (Castellucci, Kandel, 1976), возбудимые клетки изменяют свое поведение в ответ на раздражение. Учитывая, что контроль экспрессии, локализации и активности клеточных рецепторов и ионных каналов представляет собой молекулярную основу неассоциативного обучения у животных (Kandel, 2001), возникает соблазн задаться вопросом, как растения модулируют различные потоки ионов и какие элементы, придающие пластичность обучению растений.
е. Ca v , Na v ) препятствует распространению начальной деполяризации на большие расстояния. В возбудимых клетках деполяризация обеспечивает необходимую энергию для индукции открытия потенциалзависимых каналов (т. е. Ca v с, Na v с и K v с) (Bezanilla, 2008; Catterall et al., 2017). Скорость распространения, форма распространяемого потенциала и частота электрического сообщения определяются свойствами кабеля ячейки, которые определяются как геометрией каждого конкретного типа ячейки, так и набором доступных ионных каналов. У животных этот тип связи распространяется и на многоклеточный организм, когда выделившееся из данной клетки вещество оказывает влияние на постсинаптическую клетку (Gerber, Südhof, 2002; Jackson, 2006; Catterall, Few, 2008). Как описано первоначально в Aplysia Э. Канделя (Castellucci, Kandel, 1976), возбудимые клетки изменяют свое поведение в ответ на раздражение. Учитывая, что контроль экспрессии, локализации и активности клеточных рецепторов и ионных каналов представляет собой молекулярную основу неассоциативного обучения у животных (Kandel, 2001), возникает соблазн задаться вопросом, как растения модулируют различные потоки ионов и какие элементы, придающие пластичность обучению растений.
Потенциалы действия были зарегистрированы у водорослей и высших растений (Pickard, 1973; Требач и Завадски, 1985 г.; Катерия и др., 2004; Фромм и Лотнер, 2007 г.; Hegenauer и др., 2016). Как и предвидел Davies (1987), в настоящее время общепризнано, что электрическая передача сигналов играет важную роль в меж- и внутриклеточной коммуникации растений. Однако, в отличие от детальных знаний о молекулярных и клеточных механизмах, управляющих передачей электрических сигналов у животных, идентичность клеточных сенсоров и эффекторов и их точное распределение в растении до сих пор неясны (Ward et al., 2009).; Хедрих, 2012; Хедрич и др., 2016). Более того, из-за отсутствия сложной клеточной проводки, разработанной многоклеточными животными для передачи своих электрических сигналов на большие расстояния, кажется, что растения разработали архитектуру, позволяющую им формировать — или вынуждающую их адаптировать — электрическую связь по-другому. Из этого рассуждения возникают простые вопросы: как именно растения подключаются? Как электрические сигналы проходят через сотовую сеть? Как эти сигналы работают вместе, связывая зондирование окружающей среды, экспрессию генов, поглощение питательных веществ, газообмен, водный баланс, производство энергии и хранение отходов? В этом обзоре мы не стремимся ответить на такие амбициозные вопросы, а скорее рассмотрим различные элементы, которые могут способствовать генерации и распространению электрического сообщения в корнях наземных растений.
Способность ориентироваться является атрибутом животных и требует решения фундаментальных проблем, таких как (i) мультиплексный сенсорный ввод на высоких частотах, (ii) быстрая интеграция этих сигналов и (iii) точное выполнение вычисляемой команды. клеточная точность, позволяющая когерентно реагировать на тело. Одноклеточные зеленые водоросли, предки наземных растений, обладают способностями к навигации и, по случайному совпадению, представляют другой набор ионных каналов по сравнению с их потомками, экспрессируя важные элементы, важные для формирования электрического ответа возбудимых клеток у животных (Merchant et al., 2007; Wheeler). и Браунли, 2008 г.). Среди них активируемые напряжением кальциевые и натриевые каналы, каналы TRP и рианодиновый рецептор, которые отсутствуют у современных наземных растений (Wheeler and Brownlee, 2008; Ward et al., 2009).; Фромм и Лотнер, 2012 г.; Тейлор и др., 2012 г.; Ариас-Дарраз и др., 2015 г.; Эдель и др., 2017). Животным и растениям, оказавшимся в ловушке одного и того же природного мира, свойственен большой набор стрессовых факторов окружающей среды.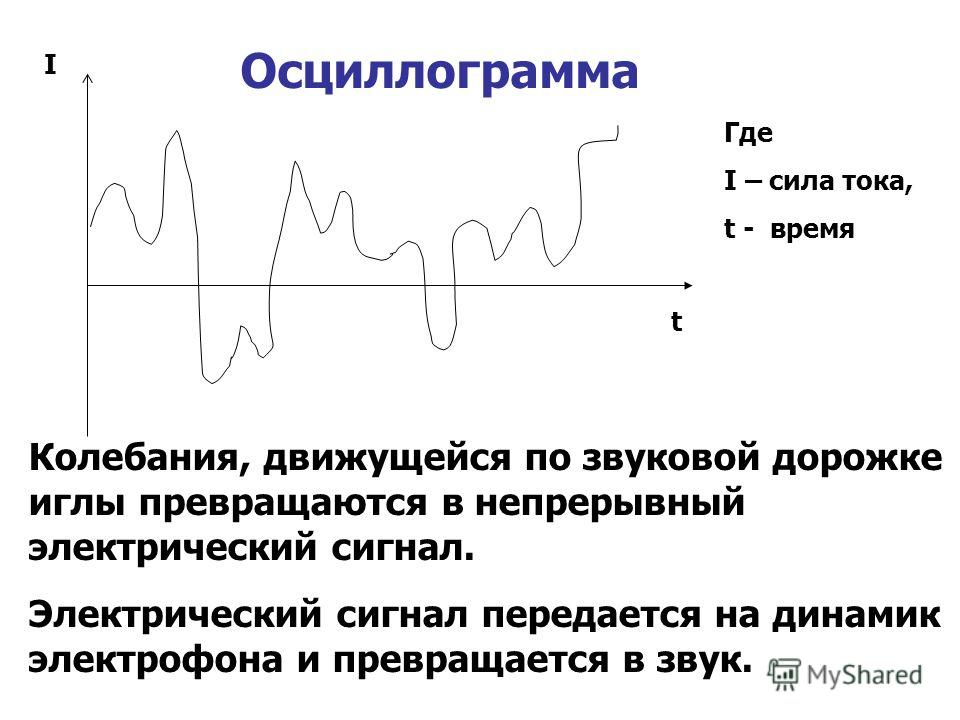 Тем не менее, они явно приняли разные пути решения основных проблем, таких как воспроизводство и самосохранение. Вероятно, поиск пищи, спаривание и потребность в удалении отходов побудили животных разработать сигнальные механизмы, настроенные на навигацию. С другой стороны, растения не только производят собственные углеводы, но и, что важно, хранят отходы. Следовательно, сидячий характер наземных растений требует вместо этого надежных механизмов адаптации. Соответственно, клеточные и молекулярные датчики постоянно снабжают растение полезной информацией об окружающей среде, которая должна распространяться по всему телу (Karban, 2015). Недавние исследования показывают, что арабидопсис эффективно организует свое трехмерное планирование для оптимизации снабжения питательными веществами (Conn et al., 2017), укрепляя идею о том, что архитектура растения контролируется механизмом управления, отвечающим за обмен между общей длиной ветвей и распределением питательных веществ. . Такой механизм должен быть связан с природой и свойствами распространения электрических сигналов, генерируемых корнями и листьями, тканями, где поглощаются минералы и вода, вырабатываются углеводы и накапливаются побочные продукты.
Тем не менее, они явно приняли разные пути решения основных проблем, таких как воспроизводство и самосохранение. Вероятно, поиск пищи, спаривание и потребность в удалении отходов побудили животных разработать сигнальные механизмы, настроенные на навигацию. С другой стороны, растения не только производят собственные углеводы, но и, что важно, хранят отходы. Следовательно, сидячий характер наземных растений требует вместо этого надежных механизмов адаптации. Соответственно, клеточные и молекулярные датчики постоянно снабжают растение полезной информацией об окружающей среде, которая должна распространяться по всему телу (Karban, 2015). Недавние исследования показывают, что арабидопсис эффективно организует свое трехмерное планирование для оптимизации снабжения питательными веществами (Conn et al., 2017), укрепляя идею о том, что архитектура растения контролируется механизмом управления, отвечающим за обмен между общей длиной ветвей и распределением питательных веществ. . Такой механизм должен быть связан с природой и свойствами распространения электрических сигналов, генерируемых корнями и листьями, тканями, где поглощаются минералы и вода, вырабатываются углеводы и накапливаются побочные продукты. Необходимы дальнейшие экспериментальные работы на неповрежденных живых растениях с использованием подходящих моделей, допускающих одновременную регистрацию электрических сигналов и изображений, для оценки влияния электрических сигналов на распределение пищи по растению (Kanchiswamy et al., 2014; Salvador-Recatalà et al., 2014; Gunsé et al., 2016; Candeo et al., 2017).
Необходимы дальнейшие экспериментальные работы на неповрежденных живых растениях с использованием подходящих моделей, допускающих одновременную регистрацию электрических сигналов и изображений, для оценки влияния электрических сигналов на распределение пищи по растению (Kanchiswamy et al., 2014; Salvador-Recatalà et al., 2014; Gunsé et al., 2016; Candeo et al., 2017).
Проводящее растение
При отсутствии не только клеточной архитектуры, но и набора ионных каналов, кодирующих электрические сообщения у животных (Ward et al., 2009; Hedrich, 2012), нет оснований предполагать, что сенсорный ввод у растений либо интегрируется, либо обрабатывается аналогичным образом. Было высказано предположение, что флоэма растения образует единый проводящий кабель, эквивалентный аксону в одном многоклеточном нейроне (Hedrich et al., 2016). Различные типы клеток, включая клетки-компаньоны и ситовидные элементы, образуют флоэму. В отличие от других типов растительных клеток, клетки ситовидных элементов не имеют разрывов в своей проницаемости из-за наличия ситовидных пластинок, обеспечивающих непрерывный транспорт растворенных веществ между различными органами растения и обеспечивающих канал с низким сопротивлением и высокой емкостью, который позволяет распространение относительно медленных электрических сигналов. Десятилетия теоретических и экспериментальных данных выдвинули концепцию, что флоэма будет основным каналом, способным электрически соединять корни и надземные ткани (Brenner et al., 2006; Fromm et al., 2013; Hedrich et al., 2016). Тем не менее, информация, обнаруженная в эпидермальных клетках корня, должна распространяться по клеточной сети коры, интегрироваться и достигать флоэмы, чтобы трансдуцироваться по всему телу растения. И наоборот, сигнал должен выйти из флоэмы, чтобы воздействовать на клетки надземной ткани. Для выполнения этой сложной задачи сосудистые растения имеют взаимосвязанное внеклеточное пространство между плазматической мембраной и клеточной стенкой (т. е. апопластическое пространство) и прямую клеточную связь через плазмодесмы (Sattelmacher and Horst, 2007; Lee, 2015). Эти особенности служат различным сигнальным функциям в растении, обеспечивая не только прохождение растворимых сигналов, но также определяя электрическую связь между клетками и модуляцию специфических сигналов, связанных с реакцией кальция, которая сочетается с обнаружением различных сигналов окружающей среды (Zebelo). et al., 2012; Nawrath et al., 2013; Lee, 2015; Choi et al., 2016; Edel et al., 2017).
У растений описаны два основных типа дальних электрических сигналов: потенциалы действия (ПД) и вариационные потенциалы (ВП) (Bose, 1907; Pickard, 1973; Fromm and Lautner, 2007, 2012). Первые индуцируются деполяризацией напряжения, проявляют пороговый потенциал, следуют принципу «все или ничего» и распространяются с постоянной скоростью и амплитудой, очень похоже на ПД, наблюдаемые в животном мире (Zawadzki et al., 1991; Jackson, 2006). ; Армстронг, 2007; Ян и др., 2016). Напротив, VP, как было показано, индуцируются быстрым увеличением внутреннего давления в ксилеме и проявляются в виде медленных волн деполяризации разного размера (Fromm and Lautner, 2007, 2012). Также была описана третья мода электрического сигнала, получившая название системных потенциалов (СП), состоящая из гиперполяризации, которая распространяется на средние расстояния (Zimmermann et al., 2009).). В то время как VP зависят от инактивации P-type H + -ATPase, SP, по-видимому, вызывается активацией помпы. Из ранних работ Бердона-Сандерсона известно, что времена нарастания ПД растений составляют порядка 0,1 с, длительность около 1 с и скорость распространения порядка нескольких сотен мм с −1 (Сандерсон, 1872; Пикард, 1973). Эти электрические сигналы различаются не только по форме и величине, но и по скорости их распространения в диапазоне от 1 до 60 мм с 9 .0035 −1 для AP до нескольких минут на сантиметр в VP. Системные потенциалы запускаются деполяризацией, не имеют характера «все или ничего», самораспространяются с постоянной скоростью около 0,5–2 мм с –1 , а их величина пропорциональна входным стимулам (Zimmermann et al. ., 2009). Интересно, что SPs напоминают рецепторные потенциалы животных, самораспространяясь одновременно по сенсорному эпителию. Лист арабидопсиса, бобов и ячменя проявляет самораспространяющуюся электрическую активность, вызванную повреждением, ограниченную связью между листьями и связанную с экспрессией генов, подобных глутаматным рецепторам (Zimmermann et al. , 2009).; Мусави и др., 2013; Сальвадор-Рекатала и др., 2014 г.; Сальвадор-Рекатала, 2016а). В этом случае тип раны, по-видимому, связан с различными типами деполяризации. Было высказано предположение, что анатомия ткани будет иметь значение для определения связи между поверхностной тканью и флоэмой (Salvador-Recatalà, 2016a).
Тем не менее было трудно систематизировать как теоретическую модель, интегрирующую электрические сигналы всего предприятия, так и экспериментальные методы изучения дальней электрической связи в конфигурации всего предприятия (Goldsworthy, 19).83; Дэвис, 1987; Петрушка и др., 1997; Фромм и Лотнер, 2007 г.; Волков, 2012; Фромм и др., 2013; Хедрич и др., 2016). Тем не менее было установлено, что различные органы растения, включая листья, стебель, цветки и корень, обладают внутренней электрической активностью (Pickard, 1973; Baldwin et al., 2006; Fromm and Lautner, 2007, 2012; Appel and Cocroft). , 2014; Engineer et al., 2015; Karban, 2015; Zhou et al., 2016). Кроме того, была описана дальняя электрическая связь между корнями, побегами и листьями (подробно рассмотрена в Pickard, 19).73; Фромм и Лотнер, 2007 г.; Циммерманн и др., 2009 г.; Хедрич и др., 2016). Подробное описание передачи электрического сигнала на корнях отсутствует, вероятно, из-за кажущейся нескоординированной природы ПД корней (Fromm and Eschrich, 1993; Fromm et al., 1997, 2013; Masi et al., 2015; Salvador-Recatalà, 2016b). ).
Картирование набора ионных каналов у арабидопсиса
Электрические свойства клеток зависят от экспрессии и контроля ионных каналов, транспортеров и насосов. Их можно модулировать различными стимулами, такими как: давление, экзогенные и эндогенные лиганды, температура, свет, напряжение мембраны и растяжение среди других. Молекулярный механизм, описывающий распространение электрических сигналов в растениях, в деталях неизвестен, но, принимая во внимание экспериментальные данные и доступную генетическую информацию, мы узнали, что растения и животные используют разные стратегии для распространения AP. В то время как животные используют чувствительный к напряжению Na + и Ca 2+ каналов для запуска деполяризации (Hodgkin and Huxley, 1952; Armstrong, 2007; Catterall et al., 2017), токсическая природа натрия заставляет растительные клетки утилизировать Cl — и Ca 2 + вместо. В то время как Ca 2+ будет вызывать деполяризацию, входя в клетку, Cl — будет делать это, выходя из клетки. Согласно профилям генной экспрессии, деполяризация растительных клеток, вероятно, управляется хлоридными каналами типа ALMT/QUAC и/или ионными каналами, обеспечивающими приток кальция, такими как двухпоровые каналы (TPC), циклические нуклеотид-управляемые каналы (CNGC) или каналы, подобные глутаматным рецепторам (GLR) (Ward et al., 2009).; Хедрих, 2012; Хедрич и др., 2016). В цепочке событий, определяющих ПД, начальный подъем Ca 2+ вызовет отток Cl — , а последующая активация потенциал-зависимых калиевых каналов, вероятно, будет участвовать в реполяризации (Schroeder et al. , 1984; Ward et al. и др., 2009; Хедрич и др., 2016).
Поскольку способность ткани генерировать электрические сигналы будет определяться набором ионных каналов, экспрессируемых в различных типах клеток, участвующих в передаче электрического сообщения, мы нанесли на карту функционально охарактеризованные каналы и транспортеры, которые ранее были связаны с электрическими сигналами. сигнализация в Arabidopsis thalina (Barbier-Brygoo et al., 2011; Hedrich, 2012) (табл. 1). Мы провели иерархический кластерный анализ, чтобы сгруппировать эти гены в соответствии с профилями экспрессии, полученными из EPlant (Waese et al., 2017). При сравнении всех соответствующих тканей на разных стадиях развития мы заметили, что разные ионные каналы представляют собой характерный паттерн экспрессии (рис. 1А).
Таблица 1 . Функциональные ионные каналы арабидопсиса.
Рисунок 1 . Картирование экспрессии ионных каналов в Arabidopsis thaliana . (A) Иерархическая кластеризация нормализованных уровней мРНК генов, кодирующих калиевые (черные), кальциевые (красные) и анионные (синие) каналы. Данные микрочипа были получены и визуализированы из браузера eFP Arabidopsis (Waese et al., 2017). Значения экспрессии для каждого гена были преобразованы в Z-баллы для всех образцов, чтобы идентифицировать тканеспецифическую экспрессию. Высокие значения Z-оценки (красный или синий) указывают на большее отклонение от средней экспрессии во всех тканях. (B) Примеры ионных каналов, демонстрирующих тканеспецифическую экспрессию.
Известно, что передача сигналов кальция важна для контроля открывания устьиц (Laanemets et al., 2013), а калиевые каналы являются критическими в фазе реполяризации (Schroeder et al., 1984). Как и ожидалось, мы наблюдали большую экспрессию каналов GLR и K v в листьях (рис. 1А, В). Хотя гены, кодирующие анионные и калиевые каналы, не показывают четкого разделения между надземной частью и корнями (рис. 1А), мы обнаружили специфические гены, экспрессия которых преобладает в корнях (VDAC1), стеблях (GLR3.2) или листьях ( GLR3.3) (рис. 1В). Эти тканеспецифичные профили экспрессии позволяют предположить, что существуют разные пути генерации и распространения электрических сигналов в растениях и что эти пути меняются во время развития. При более подробном наблюдении два специфичных для корней анионных канала, SLAH4 и VDAC1, показали различный характер экспрессии в разных типах клеток (рис. 2). Потенциал-зависимый анионный канал VDAC1 сильно выражен вдоль ткани корня. Для сравнения, экспрессия в меристематической зоне выше, чем в корневых волосках (рис. 2). Аналогично, H + -АТФаза экспрессируется почти во всех типах клеток корня. Напротив, медленный хлоридный канал проводимости SLAH4 проявлял большую экспрессию во внутренних тканях корня, таких как перицикл и кора, важных физических барьерах на пути к флоэме (Nawrath et al., 2013). В то время как экспрессия электрогенного переносчика нитратов NRT1. 1 наблюдается преимущественно в корневых волосках и на флоэме ближе к стеблю, вакуолярный канал TPC1 заметно выражен в корневых волосках зоны созревания. Интересно, что ни один из этих мембранных белков не проявлял заметной экспрессии во флоэме вдоль корня (рис. 2).
Рисунок 2 . Сравнительные профили экспрессии при разрешении анионных каналов и генов клеточного типа, связанных с поглощением нитратов в корне арабидопсиса. Данные микрочипов были получены и визуализированы с помощью браузера Arabidopsis eFP (Waese et al., 2017).
Приток кальция в ответ на внешние раздражители кажется критическим для генерации электрического сигнала. Около 50 различных ионных каналов связаны с притоком Ca 2+ в наземные растения. Этот большой набор кальциевых каналов разделен на пять разных семейств: CNGC, GLR, TPC1, каналы, связанные с осмотическим ответом (OSCA), и механочувствительные кальциевые каналы (MCA) (Kurusu et al., 2012; Chin et al., 2013; Morgan and Galione, 2014; Edel et al. , 2017) (табл. 2). Из этих каналов CNGC14, CNGC19, GluR2.1 и OSCA1.4, по-видимому, преимущественно экспрессируются в ткани корня (рис. 3А). Экспрессия этих каналов также была разной. CNGC14 в значительной степени экспрессируется в эпителии вблизи зоны меристемы и в меньшей степени в зоне созревания. Напротив, вакуолярный канал CNGC19 концентрируется в эндотелии и флоэме. С другой стороны, GLR2.1, управляемые лигандом, и осмотически связанные каналы OSCA1.4 предпочтительно экспрессируются в корневых волосках. В то время как экспрессия GLR2.1 в эпителиальной ткани несколько уменьшается от зоны меристемы к зоне созревания, OSCA1.4 предпочтительно экспрессируется в зоне созревания и флоэме (рис. 3В).
Таблица 2 . Кальциевые каналы арабидопсиса.
Рисунок 3 . Картирование экспрессии кальциевых каналов у Arabidopsis thaliana . (A) Тепловая карта и кластерные дендрограммы экспрессии кальциевых каналов среди типов тканей. Образцы тканей представлены в столбцах, а гены — в строках. Значения экспрессии для каждого гена были преобразованы в Z-показатели для всех образцов, чтобы определить специфичную для корней экспрессию, которая обозначена черными прямоугольниками. (B) Кальциевые каналы с характерной для корня экспрессией.
Обработка нитратами растений, испытывающих дефицит азота, вызывает временную деполяризацию плазматической мембраны (Meharg and Blatt, 1995; Wang and Crawford, 1996; Wang et al., 1998). Точно так же недавно сообщалось, что лечение нитратами вызывает увеличение внутриклеточного кальция, что инициирует сигнальный путь нитратов (Liu et al., 2017). Более того, генетические данные указывают на то, что повышение уровня внутриклеточного кальция связано с НЗТ1.1. (Риверас и др., 2015). Учитывая наблюдаемый профиль экспрессии, мы можем предположить, что деполяризация эпителиальных клеток, вызванная поглощением нитратов, вызванная увеличением активности переносчика нитратов NRT1. 1, будет запускать приток кальция через OSCA1.4 и/или GLR2.1. , дополнительно активируя проводимость кальция или хлорида, зависящую от напряжения (например, SLAh4 или VDAC1), что позволяет распространять электрический сигнал вдоль коры к флоэме (рис. 4).
Рисунок 4 . Схематическое изображение корня растения, показывающее различные пути распространения электрических сигналов. Различное проводящее состояние плазмодесмы A, B и C будет модулировать сигнал, обнаруженный в корневом волоске, на пути к ситовидной трубке. На верхней левой панели показаны проводимости, которые могут быть связаны с деполяризацией, вызванной нитратами. Увеличение содержания кальция в эпителиальных клетках модулирует активность плазмодесмы. Как уровни кальция, так и физические изменения плазмодесмы могут модулировать активность ионных каналов, выраженных в этой области. На верхней правой панели изображены цитоплазматический и ретикулярный пути для ионов, оба из которых могут быть модулированы взаимодействием с закрепляющими белками в месте контакта поры.
Сотовая связь как часть передачи электрических сигналов растениями
Необходима объединенная модель, связывающая чувствительность растений и контролируемое поведение, чтобы объяснить, могут ли стимулы, обнаруженные на границах тела растения (например, корневые волоски, эпителиальные клетки листьев), преобразовывать электрическую информацию к кабелю флоэмы, как сигнал далее интегрируется и, наконец, как электрический сигнал выходит из флоэмы, достигая эффекторной ткани, расположенной в отдаленном эпителии (например, замыкающие клетки в устьице листа). В то время как дифференциальное выражение набора ионных каналов важно для определения возбудимости, взаимосвязанная клеточная архитектура растений будет иметь решающее значение для управления амплитудой и свойствами распространения в трехмерном пространстве. Модуляция этих элементов в ответ на повторяющиеся, острые или хронические сенсорные воздействия может придать пластичность реакции растения и позволит как обучаться, так и адаптироваться без необходимости в «мозгоподобном» интеграторе или когнитивном поведении, как это предлагается в литературе. Балуска и др., 2004).
Одним из основных структурных различий между клетками животных и растений является наличие клеточной стенки. Клеточная стенка растений создает необычную внеклеточную среду, известную как апопласт, который представляет собой физический/химический барьер, участвующий в путях межклеточной коммуникации, включая передачу электрических сигналов на большие расстояния (Sattelmacher, 2001; Fromm and Lautner, 2007; Choi et al. ., 2017). У растений внеклеточная концентрация ионов соответствует концентрации апопластных ионов, которые строго регулируются. Ионный состав апопласта изменчив и зависит как от внутренних факторов, таких как ткань или стадия развития, так и от внешних факторов, таких как пищевой стресс (López-Millán et al., 2001). Ионы в апопластическом компартменте обычно находятся в низких концентрациях, состоящих преимущественно из неорганических катионов и анионов, таких как K + , Ca 2+ , Mg 2+ , Cl — , NO3- и PO43- (Gabriel and Kesselmeier, 1999). Концентрации апопластных и внутриклеточных ионов вместе с ионной проницаемостью будут определять потенциал покоя эпидермальной клетки корня и, как сообщается, составляет около -120 мВ, отрицательный внутри (Fromm and Eschrich, 1993). Биохимические свойства клеточной стенки будут определять в первую очередь апопластическое пространство и проницаемость для питательных веществ. У взрослого растения высокое содержание лигнина или суберина в клеточных стенках значительно снижает их проницаемость (Nawrath et al., 2013) и, таким образом, представляет собой плотный барьер для свободной диффузии питательных веществ в ткани растения и из них. Напротив, структура и состав клеточных стенок изменяются в процессе развития корня (Somssich et al., 2016). Меристематическая зона содержит молодые клетки, локализованные ближе к кончику корня, а более старые клетки локализованы у основания корня ближе к стеблю. Клеточные стенки меристематической зоны тонкие и более проницаемые из-за более высокой митотической активности (Baluska et al. , 19).96). Клетки из зоны дифференцировки более ригидны за счет накопления лигнина, связанного с развитием вторичных клеточных стенок, что придает стенкам дополнительную прочность и делает их водонепроницаемыми Somssich et al. (2016). Следовательно, в первичных корнях существует продольный градиент проницаемости, определяемый степенью дифференцировки эпидермальных клеток.
Равновесную концентрацию ионов можно рассчитать по уравнению Нернста вида [X] out /[X] в = exp(V m ZF/RT), где [X] out и [X] в – внешняя и внутренняя концентрации иона X, V m – напряжение на мембране, Z — валентность проницаемого иона, F — постоянная Фарадея, R — газовая постоянная, T — абсолютная температура. Из этого уравнения ясно, что изменения внеклеточной концентрации проницаемого заряженного растворенного вещества будут влиять на мембранный потенциал в состоянии покоя. Фактически, ранние исследования показали, что как пик потенциала действия, так и величина входящего тока зависят от концентрации апопластного кальция (Hope, 19). 61а,б; Хоуп и Финдли, 1964). С другой стороны, активность Н + -АТФазы поддерживает рН апопласта, обычно кислый (4,7–5) (Sattelmacher and Horst, 2007). Поскольку мембранный потенциал покоя в растительных клетках, вероятно, определяется транспортом протонов, следует ожидать высокой чувствительности к изменениям активности протонной помпы (Brault et al., 2004). Учитывая, что апопласт занимает относительно небольшой объем растительных тканей, составляя менее 5% в случае листьев (López-Millan et al., 2001), разумно ожидать, что изменения окружающей среды могут вызвать быстрое подщелачивание апопласта, что в свою очередь вызовет локальную деполяризацию (Grams et al., 2009).). Анионные питательные вещества, такие как нитраты, сульфаты и фосфаты, усваиваются пассивно посредством электрогенного котранспорта, которому помогает протонный градиент (Ullrich-Eberius et al., 1981; Muchhal and Raghothama, 1999). Точно так же рН апопласта будет чувствителен к активности переносчиков фосфатов и нитратов (Amtmann et al. , 1999). Соответственно, острая обработка нитратами растений, испытывающих недостаток азота, вызывает временную деполяризацию плазматической мембраны (Meharg and Blatt, 1995; Wang and Crawford, 19).96). Экспериментальные данные свидетельствуют о том, что при поглощении фосфата котранспортируются 2–4 протона (Sakano, 1990). В случае нитрата стехиометрия котранспорта с протонами была рассчитана как 2:1 (McClure et al., 1990; Glass et al., 1992; Wang and Crawford, 1996). Принимая во внимание экспериментальные данные in-vitro исследований с мембранными везикулами, а также генетические анализы, такие как комплементация дрожжей, наиболее вероятная стехиометрия для котранспорта протонов/сульфатов будет 3:1 (Buchner et al., 2004).
Корень – это первый орган, который вступает в контакт с водой и питательными веществами. Поэтому, когда растения лишены питательных веществ, корень является основным местом для обнаружения. Сравнительный транскриптомный анализ образцов корней и побегов показал, что реакция растения на нитраты инициируется в корне (Wang et al. , 2003). Растения должны интегрировать информацию из внешней среды и сопоставлять ее со своим пищевым статусом. Фактически, сообщалось, что баланс между азотом и углеродом важен для контроля усвоения азота (Zheng, 2009).). Следовательно, растениям требуется эффективная система связи между местом восприятия и поглощения питательных веществ (т. е. корнями) и метаболическим центром, где производится углерод (т. е. листьями), чтобы адекватно реагировать на изменения доступности питательных веществ. Эпидермальные клетки корня растения обладают более высокой проницаемостью через клеточную стенку, заставляя электрогенные переносчики на плазматической мембране сталкиваться с высокими концентрациями питательных веществ, когда они растворяются в почве, окружающей корень. Поглощение любого из этих питательных веществ вызовет быструю и преходящую деполяризацию мембраны эпителиальной клетки в корне (Dunlop and Gradiner, 19).93; Мехарг и Блатт, 1995). Кроме того, диффузия веществ внутри апопласта корня внутренне ограничена полосой Каспари, гидрофобной пропиткой первичной клеточной стенки лигнином, которая запечатывает внеклеточное пространство энтодермальных клеток, заставляя ионы, питательные вещества и воду проходить через первичную клеточную стенку. плазматическая мембрана эндотелиальных клеток (Nawrath et al., 2013; von Wangenheim et al., 2017) (рис. 4).
В дополнение к пути апопластной коммуникации во внеклеточном пространстве цитоплазма растительных клеток может быть внутренне связана межклеточными соединениями, известными как плазмодесма (Lee, 2015; Kitagawa and Jackson, 2017). Межклеточная коммуникация тканей корня была продемонстрирована быстрой диффузией молекул флуоресцентных индикаторов (например, иодида пропидия) через плазмодесмы во внутренние слои ткани корня (Nawrath et al., 2013). Показано, что недифференцированные клетки меристемы апикального корня и зоны растяжения связаны с красителем и, следовательно, симпластически связаны через плазмодесмы (Duckett et al., 19).94). Ионы и более крупные молекулы могут свободно диффундировать через поры от одной клетки к другой, что делает эти клеточные структуры центром передачи сигналов через ткань коры (Burch-Smith and Zambryski, 2012; Lee, 2015). Более того, сложность плазмодесм подчеркивается наличием эндоплазматического ретикулума (ЭР), проходящего через поры, что обеспечивает вторичный и, вероятно, более избирательный путь коммуникации между соседними клетками. Известно, что разные липиды и каллоза , растворимый белок, способный блокировать цитоплазматический путь, регулировать проникновение через плазмодесмы (Tilsner et al., 2016). Более того, сообщалось, что повышение цитоплазматического кальция способствует закрытию цитоплазматического пути поры (Lee, 2015). Необходимы дополнительные данные, чтобы определить вклад этих трубок ЭР в распространение как кальция, так и электрических сигналов через кору корня.
У растений прямая связь между ЭР и электрической активностью плазматической мембраны не связана с молекулой стромального взаимодействия 1 (STIM1) или кальциевыми каналами Orai1, как в клетках животных. Белки, родственные STIM, были утрачены на уровне одноклеточных водорослей, а родственники Orai присутствуют только до голосеменных (Edel et al., 2017). Тем не менее было высказано предположение, что закрепляющие белки, элементы цитоскелета и липиды могут модулировать локализацию и активность мембранных белков в месте контакта между ЭР и плазматической мембраной (Lee, 2015). Таким образом, дифференциальная экспрессия и модуляция специфического набора кальций-чувствительных ионных каналов, окклюзия плазмодесм или ремоделирование формы пор в месте контакта могут обеспечивать усиление или подавление электрического сигнала, распространяющегося через кору (рис. 4).
Дифференцировка эпидермальных клеток корней у Arabidopsis прогрессивно редуцирует эти цитоплазматические связи таким образом, что на последней стадии их развития они становятся симпластически разъединенными (Duckett et al., 1994). Наоборот, клетки эпидермиса гипокотиля симпластически связаны друг с другом независимо от состояния их развития (Duckett et al., 1994). Более ранние доказательства электрической связи были предоставлены Спансвиком и Костертоном, которые показали, что при введении тока в клетку многоклеточной водоросли Nitella сигнал можно было проследить на несколько клеток дальше от места введения (Спансвик и Костертон, 19).67).
Недавние эксперименты по визуализации кальция с использованием генетически закодированного логометрического индикатора кальция (т. е. Y-Cameleon 3.6), выраженного в живом арабидопсисе, показали, как спонтанная реакция, возникающая в корневом волоске, распространяется по ткани корня четко определенными «участками» ( Кандео и др., 2017). Хотя авторы не прокомментировали этот шаблонный ответ, по аналогии с сенсорными рецепторами у животных возникает соблазн интерпретировать такие показания как физическое измерение «9».0021 сенсорное поле », которые соответствуют определенной эпителиальной клетке или их группе. После стимуляции клетки эпидермиса корня генерируется электрический сигнал; в приведенном выше примере приток кальция может быть напрямую связан с деполяризацией клеток. Генерируемый электрический сигнал может передаваться через плазмодесмы к соседним связанным клеткам в коре корня. Как только сигнал достигает ситовидной трубки с низким сопротивлением во флоэме, он распространяется по всему растению (рис. 4). Тогда возникает вопрос, интегрирует ли флоэма множественные сигналы, поступающие от отдельных рецепторных полей, рассредоточенных по эпителию корня, и как сеть межклеточных связей может обеспечивать пластичность, модулируя распространение электрического сообщения, контролируя локализацию и активность плазмодесм. (Рисунок 4). Более того, эксперименты по диффузии с флуоресцентными красителями показали, что сообщение эпидермальных клеток гипокотиля заканчивается у основания стебля (Duckett et al., 19).94). Следовательно, вполне вероятно, что эпидермальные клетки гипокотиля и корня электрически не связаны, что делает ситовидные трубки единственным возможным путем распространения AP от корня к побегу. Все эти физические барьеры создают узлы резистивных элементов, полезных для фильтрации сигналов не только на выходе из корневой ткани, но также внутри и вне ветвей, отходящих от ствола.
Было высказано предположение, что растения могут интегрировать информацию через структуру «, похожую на мозг », расположенную в верхушке корня (Baluska et al., 2004; Brenner et al., 2006). Однако поразительные особенности мозга животных обусловлены не только тем фактом, что нейроны образуют синапсы, но и способностью создавать сложные логические схемы на основе клеток. Один нейрон может содержать сотни клеточно-специфических синапсов, каждый из которых способен к пластической адаптации. В отсутствие такого рода клеточного взаимодействия маловероятно, чтобы интеграция электрической информации во всем растении привела к когнитивной обработке сенсорной информации. Более того, недавние исследования одноклеточных организмов, таких как слизевики и бактерии, показывают, что нет необходимости в центральной нервной системе для создания интеллектуальных решений (Nakagaki, 2001; Kotula et al., 2014).
Создавая очень упрощенную картину электрической связи растений, мы видим, что она состоит из очень неэффективного кабеля, окруженного эпителиальной тканью, образованной поглощающими и возбудимыми клетками. В такой модели эпителиальные волосковые клетки корня могут функционировать как шипы в нейроне, передавая сигналы, генерируемые внешними стимулами, в цепь коры, где они обрабатываются до достижения флоэмы. В то же время переход от корня к стеблю может обеспечивать дополнительный фильтр, действующий как макроскопическая версия шейки позвоночника, модулирующая транзит дальних электрических сигналов. Количество электрически связанных клеток в коре определяет размер конденсатора цепи и обрисовывает в общих чертах электрические свойства путей, ведущих к флоэме. Присутствие гипотетических «синапсоподобных» структур, основанных на актине, может способствовать формированию сети межклеточных соединений (Baluška et al., 2005). В этом контексте системные растворимые сигналы, такие как ауксины, могут также играть важную роль в регуляции межклеточных соединений, например, путем взаимодействия с плазмодесмами (Baluska et al., 2004; Brenner et al., 2006). Кроме того, сенсорный ввод может также влиять на основные свойства кабеля вдоль канала флоэмы, формируя и фильтруя распространяющиеся потенциалы действия. Природа физических/молекулярных барьеров (например, переходы коры корня во флоэму и эпителия флоэмы в лист) будет иметь большое значение для понимания интеграции сенсорной информации у растений.
Определение питательных веществ и передача электрических сигналов дальнего действия
Растения приобретают необходимые химические компоненты для синтеза биомолекул из минералов, присутствующих в почве (Maathuis, 2009). Фосфор, азот, сера, калий, магний и кальций являются питательными веществами, которые требуются высшим растениям в больших количествах (Kirkby, 2011). Следовательно, растения должны постоянно получать эти питательные вещества, чтобы обеспечить надлежащий рост и развитие (Maathuis, 2009). В отличие от гетеротрофных организмов, растения в основном получали питательные вещества в неорганической форме с помощью специфических переносчиков, локализованных в корнях. Экспрессия этих переносчиков питательных веществ, а также их активность регулируются доступностью питательных веществ, метаболизмом и факторами окружающей среды (Giehl and von Wirén, 2014). Недостаток одного из этих необходимых питательных веществ оказывает прямое влияние на рост и развитие растений, особенно в случае корневой ткани (Gruber et al., 2013). Следовательно, растения разработали сложные регуляторные системы для обеспечения поглощения этих неорганических питательных веществ (Schachtman and Shin, 2007). Фактически, реакция на голодание по питательным веществам включает сложные сигнальные сети, включая сенсорные белки (Ho et al. , 2009).), факторы транскрипции (Rubio et al., 2001), микроРНК (Vidal et al., 2010), пептиды (Ohkubo et al., 2017) и фитогормоны (Kiba et al., 2011). Интересно, что эти сигнальные пути не только запускают краткосрочные ответы, включающие метаболические корректировки и/или регуляцию переносчиков питательных веществ, но также вызывают модификации архитектуры корневой системы.
Среди макроэлементов фосфор и азот оказывают большее влияние на архитектуру корня по сравнению с другими (Gruber et al., 2013). Фосфатное голодание сильно индуцирует развитие корневых волосков (Péret et al., 2011), тогда как добавление нитратов вызывает увеличение плотности корневых волосков (Canales et al., 2017). Кроме того, дефицит фосфатов вызывает значительное снижение роста первичных корней и стимулирует развитие боковых корней (Shahzad and Amtmann, 2017). Напротив, более низкая доступность нитратов увеличивает рост первичных корней и замедляет развитие боковых корней. Эти противоположные эффекты на клеточную архитектуру корня предполагают наличие разных сигнальных путей для нитратов и фосфатов, вероятно, связанных с электрическими сигналами разной природы, которые не обязательно распространяются одинаковым образом или генерируются в эпителиальных клетках одного и того же типа.
Заключение
Внутриклеточные кальциевые вариации в клетках корневых волосков эпидермиса растений генерируются механизмом, включающим локальные вариации мембранного потенциала, вызванные электрогенным транспортом или прямой активацией растительных рецепторов внеклеточными лигандами. Эти локальные вариации мембранного напряжения будут обеспечивать усиление входного сигнала за счет увеличения проницаемости эпителиальных клеток для кальция и дальнейшего запуска потенциалов действия за счет увеличения проницаемости хлоридных проводимостей с последующей реполяризацией, управляемой калием/протонами. Эти электрические сигналы первоначально распространяются через клетки коры с помощью строго регулируемых сетей плазмодесм и, используя континуум апопластического пространства, достигают ситовидной трубки флоэмы, где они получают доступ для передачи по всему телу растения. Физические барьеры, распределение ионных каналов и межклеточная связь в корне являются критическими аспектами, которые формируют электрические сигналы, генерируемые в сенсорной ткани. Эти элементы, находящиеся под клеточным контролем, могут обеспечивать пластичность реакции растений. Роль плазмодесм в распространении и тонкой настройке электрических сигналов в сенсорном эпителии растений является фундаментальной темой, которой несколько пренебрегают и, очевидно, она требует большего внимания.
Вклад автора
SB, JC и CH-V написали статью. SB и JC подготовили рисунки.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
MiNICAD — это ядро тысячелетия, поддерживаемое Iniciativa Científica Milenio, Министерство экономики, развития и туризма, Чили. Anillo Científico ACT-1401 поддерживает SB. SB является частью программы CISNe-UACh и UACh по клеточной биологии. JC поддерживается грантом FONDECYT 11150070. Институт интегративных систем и синтетической биологии тысячелетия поддерживается Министерством экономики, развития и туризма Чили «Iniciativa Científica Milenio». Мы благодарим Шарлотту К. Коленсо за ее комментарии к этой рукописи.
Ссылки
Амтманн, А., Джелитто, Т.С., и Сандерс, Д. (1999). K+-селективные каналы внутреннего выпрямления и апопластический pH в корнях ячменя. Завод физиол. 120, 331–338. doi: 10.1104/pp.120.1.331
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Аппель, Х.М., и Кокрофт, Р.Б. (2014). Растения реагируют на колебания листьев, вызванные жеванием травоядными насекомыми. Экология 175, 1257–1266. doi: 10.1007/s00442-014-2995-6
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Ариас-Даррас Л., Кабесас Д., Коленсо С.К., Алегрия-Аркос М., Браво-Морага Ф., Варас-Конча И. и др. (2015). Ионный канал временного рецепторного потенциала у хламидомонады имеет общие ключевые характеристики с TRP-каналами, связанными с сенсорной трансдукцией, у млекопитающих. Растительная клетка 27, 177–188. doi: 10.1105/tpc.114.131862
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Армстронг, CM (2007). Жизнь среди аксонов. год. Преподобный Физиол. 69, 1–18. doi: 10.1146/annurev.physiol.69.120205.124448
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Армстронг, CM (2015). Срок службы упаковки: происхождение ионоселективных каналов. Биофиз. Дж. 109, 173–177. doi: 10.1016/j.bpj.2015.06.012
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Balagué, C., Lin, B., Alcon, C., Flottes, G., Malmström, S., Köhler, C., et al. (2003). HLM1, важный сигнальный компонент реакции гиперчувствительности, является членом семейства ионных каналов, управляемых циклическими нуклеотидами. Растительная клетка 15, 365–379. doi: 10.1105/tpc.006999
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Болдуин И. Т., Халичке Р., Пашольд А., фон Даль К. К. и Престон К. А. (2006). Летучие сигналы во взаимодействиях между растениями: «говорящие деревья» в эпоху геномики. Наука 311, 812–815. doi: 10.1126/science.1118446
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Балуска Ф., Манкузо С., Фолькманн Д. и Барлоу П. (2004). Верхушки корней как командные центры растений: уникальный «мозгоподобный» статус переходной зоны верхушек корней. Биология 59, 7–19.
Google Scholar
Балуска Ф., Фолькманн Д. и Барлоу П. В. (1996). Специализированные зоны развития в корнях: взгляд с клеточного уровня. Завод физиол. 112, 3–4.
Реферат PubMed | Google Scholar
Балушка Ф., Фолькманн Д. и Мензель Д. (2005). Синапсы растений: домены на основе актина для межклеточной коммуникации. Trends Plant Sci. 10, 106–111. doi: 10.1016/j.tplants.2005.01.002
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Barbier-Brygoo, H., De Angeli, A., Filleur, S., Frachisse, J.-M., Gambale, F., Thomine, S., et al. (2011). Анионные каналы/транспортеры в растениях: от молекулярных оснований до регуляторных сетей. год. Преподобный завод биол. 62, 25–51. doi: 10.1146/annurev-arplant-042110-103741
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Беккер Д., Гейгер Д., Дункель М., Роллер А., Бертл А., Латц А. и др. (2004). АтТПК4, 9Канал 0021 Arabidopsis с тандемными порами K + , предназначенный для контроля напряжения пыльцевой мембраны в зависимости от pH и Ca 2+ . Проц. Натл. акад. науч. США . 101, 15621–15626. doi: 10.1073/pnas.0401502101
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Беллоно, Н. В., Байрер, Дж. Р., Лейтч, Д. Б., Брайерли, С. М., Ингрэм, Х. А., Джулиус, Д., и др. (2017). Энтерохромаффинные клетки представляют собой хемосенсоры кишечника, которые соединяются с сенсорными нервными путями. Статьи энтерохромаффинных клеток являются хемосенсорами кишечника, которые связываются с сенсорными нервными путями. Сотовый 170, 1–14. doi: 10.1016/j.cell.2017.05.034
CrossRef Полный текст | Google Scholar
Берриер К. , Пейроннет Р., Беттон Дж. М., Эфритихин Г., Барбье-Бригу Х., Фрашисс Дж. М. и др. (2015). Канальные характеристики VDAC-3 от Arabidopsis thaliana . Биохим. Биофиз. Рез. Коммуна . 459, 24–28. doi: 10.1016/j.bbrc.2015.02.034
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Боймер, Дж., и Клеверс, Х. (2017). Как кишечник чувствует, пахнет и разговаривает. Сотовый 170, 10–11. doi: 10.1016/j.cell.2017.06.023
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Безанилья, Ф. (2008). Как мембранные белки чувствуют напряжение. Нац. Преподобный Мол. Клеточная биол. 9, 323–332. doi: 10.1038/nrm2376
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Бозе, Дж. К. (1907). Сравнительная электрофизиология, физико-физиологическое исследование. Лондон: Лонгманс.
Бойд, Калифорния (2008). Факты, фантазии и забавы в эпителиальной физиологии. Экспл. Физиол. 93, 303–314. doi: 10.1113/expphysiol.2007.037523
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Браулт М., Амиар З., Пеннарун А.-М., Монестез М., Чжан З., Корнел Д. и др. (2004). Деполяризация плазматической мембраны, индуцированная абсцизовой кислотой в суспензионных клетках арабидопсиса, включает снижение перекачки протонов в дополнение к активации анионных каналов, оба из которых зависят от Са2+. Завод физиол. 135, 231–243. дои: 10.1104/стр.103.039255
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Бреннер Э. Д., Штальберг Р., Манкузо С., Виванко Дж., Балушка Ф. и Ван Волкенбург Э. (2006). Нейробиология растений: комплексный взгляд на сигнализацию растений. Trends Plant Sci. 11, 413–419. doi: 10.1016/j.tplants.2006.06.009
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Бюхнер П., Такахаши Х. и Хоксфорд М. Дж. (2004). Транспортеры сульфатов растений: координация поглощения, внутриклеточного и дальнего транспорта. Дж. Эксп. Бот. 55, 1765–1773 гг. doi: 10.1093/jxb/erh306
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Берч-Смит, Т. М., и Замбрыски, П. К. (2012). Сдвиг парадигмы Plasmodesmata: регулирование извне против внутреннего. год. Преподобный завод биол. 63, 239–260. doi: 10.1146/annurev-arplant-042811-105453
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Каналес Дж., Контрерас-Лопес О., Альварес Дж. М. и Гутьеррес Р. А. (2017). Индукция нитратами плотности корневых волосков опосредована транскрипционными факторами TGA1/TGA4 и CPC в Arabidopsis thaliana . Завод J. 92, 305–316. doi: 10.1111/TPJ.13656
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кандео А., Доккула Ф. Г., Валентини Г., Басси А. и Коста А. (2017). Световая листовая флуоресцентная микроскопия количественно определяет колебания кальция в корневых волосках Arabidopsis thaliana . Физиология клеток растений. 58, 1–12. doi: 10.1093/pcp/pcx045
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Кастеллуччи, В., и Кандель, Э. (1976). Пресинаптическая фасилитация как механизм поведенческой сенсибилизации при аплизии. Наука 194, 1176–1178. doi: 10.1126/science.11560
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Catterall, WA, and Few, AP (2008). Регуляция кальциевых каналов и пресинаптическая пластичность. Нейрон 59, 882–901. doi: 10.1016/j.neuron.2008.09.005
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Catterall, W. A., Wisedchaisri, G., and Zheng, N. (2017). Химическая основа электрической сигнализации. Нац. хим. биол. 13, 455–463. doi: 10.1038/nCHEMBIO.2353
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чин К., Дефалько Т. А., Модер В. и Йошиока К. (2013). Ионные каналы AtCNGC2 и AtCNGC4, управляемые циклическими нуклеотидами арабидопсиса, работают по одному и тому же сигнальному пути, регулируя защиту от патогенов и цветочный переход. Завод физиол. 163, 611–624. doi: 10.1104/pp.113.225680
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чой В.-Г., Хиллари Р., Суонсон С.Дж., Ким С.-Х. и Гилрой С. (2016). Быстрая электрическая и кальциевая передача сигналов на большие расстояния в растениях. год. Преподобный завод биол. 67, 287–307. doi: 10.1146/annurev-arplant-043015-112130
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чой В.-Г., Миллер Г., Уоллес И., Харпер Дж., Миттлер Р. и Гилрой С. (2017). Организация быстрой передачи сигналов на большие расстояния у растений с Ca 2+ , АФК и электрические сигналы. Завод J. 90, 698–707. doi: 10.1111/tpj.13492
Полный текст CrossRef | Google Scholar
Клемменсен, К., Мюллер, Т. Д., Вудс, С. К., Берту, Х., Сили, Р. Дж., и Чеп, М. Х. (2017). Взаимодействие кишечника и мозга в метаболическом контроле. Сотовый 168, 758–774. doi: 10.1016/j.cell.2017. 01.025
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Коул, К.С., и Кертис, Х.Дж. (1939). Электрический импеданс гигантского аксона кальмара во время активности. J. General Physiol. 22, 649–670. doi: 10.1085/jgp.22.5.649
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Конн А., Педмале У. В., Чори Дж., Стивенс К. Ф. и Навлаха С. (2017). Статистическое описание строения побегов растений. Курс. биол. 27, 2078.e3–2088.e3. doi: 10.1016/j.cub.2017.06.009
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Дарвин, К. (1897 г.). Насекомоядные растения . изд. Д. Эпплтон. Нью-Йорк, штат Нью-Йорк: Дж. Мюррей.
Дэвис, Э. (1987). Потенциалы действия как многофункциональные сигналы у растений: объединяющая гипотеза для объяснения явно несопоставимых реакций на рану. Окружающая среда растительных клеток. 10, 623–631.
Google Scholar
Де Анджели А. , Моначелло Д., Эфритихин Г., Фрашисс Дж. М., Томин С., Гамбаль Ф. и др. (2006). Нитрат/протонный антипортер AtCLCa опосредует накопление нитратов в вакуолях растений. Природа . 442, 939–942. doi: 10.1038/nature05013
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Де Анджели А., Чжан Дж., Мейер С. и Мартинойя Э. (2013). AtALMT9 представляет собой активируемый малатом вакуолярный хлоридный канал, необходимый для открытия устьиц у Arabidopsis. Нац Коммуна . 4, 1804. doi: 10.1038/ncomms2815
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Дакетт К.М., Опарка К.Дж., Прайор Д.А.М., Долан Л. и Робертс К. (1994). Связывание красителя в эпидермисе корней арабидопсиса постепенно снижается в процессе развития. Разработка 3255, 3247–3255.
Google Scholar
Данлоп, Дж., и Градинер, С. (1993). Поглощение фосфатов, экструзия протонов и мембранные электропотенциалы фосфордефицитных Trifolium repens L. J. Exp. Бот. 44, 1801–1808 гг. doi: 10.1093/jxb/44.12.1801
CrossRef Полный текст | Google Scholar
Эдель, К. Х., Маршадье, Э., Браунли, К., Кудла, Дж., и Хетерингтон, А. М. (2017). Эволюция передачи сигналов на основе кальция у растений. Курс. биол. 27, Р667–Р679. doi: 10.1016/j.cub.2017.05.020
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Engineer, CB, Hashimoto-Sugimoto, M., Negi, J., Israelsson-Nordström, M., Azoulay-Shemer, T., Rappel, W. J., et al. (2015). Чувствительность к СО2 и регуляция устьичной проводимости с помощью СО2: достижения и открытые вопросы. Trends Plant Sci. 21, 16–30. doi: 10.1016/j.tplants.2015.08.014
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Финкельштейн, А. (1976). Водо- и неэлектролитная проницаемость липидных бислойных мембран. J. Общая физиол. 68, 127–135. doi: 10.1085/jgp.68.2.127
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Фрингс, С. (2009). Первичные процессы в сенсорных клетках: современные достижения. Дж. Комп. Физиол. Нейроэтол. Сенсор Нейронный. Поведение Физиол. 195, 1–19. doi: 10.1007/s00359-008-0389-0
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Фромм Дж. и Эшрих В. (1993). Электрические сигналы, испускаемые корнями ивы ( Salix viminalis L.), изменяют транспирацию и фотосинтез. J. Физиол растений. 141, 673–680. doi: 10.1016/S0176-1617(11)81573-7
CrossRef Full Text | Google Scholar
Фромм Дж., Хаджирезаи М.-Р., Беккер В.К. и Лотнер С. (2013). Электрическая передача сигналов по флоэме и ее физиологические реакции в листьях кукурузы. Фронт. Растениевод. 4:239. doi: 10.3389/fpls.2013.00239
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Фромм Дж. и Лотнер С. (2007). Электрические сигналы и их физиологическое значение в растениях. Окружающая среда растительных клеток. 30, 249–257. doi: 10.1111/j.1365-3040.2006.01614.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Фромм Дж. и Лотнер С. (2012). «Электрофизиология растений: сигналы и ответы», в Электрофизиология растений , изд. А. Г. Волкова (Берлин; Гейдельберг: Springer-Verlag), 1–377.
Фромм, Дж., Мейер, А.Дж., и Вайсензел, М.Х. (1997). Абсцизовая кислота и спермин влияют на рост, мембранный потенциал и эндогенные ионные токи корней ивы ( Salix viminalis ). Физиол. Завод. 99, 529–537. doi: 10.1034/j.1399-3054.1997.9
.xПолный текст CrossRef | Google Scholar
Габриэль Р. и Кессельмайер Дж. (1999). Концентрации апопластных растворов органических кислот и минеральных питательных веществ в листьях некоторых буковых. Физиология клеток растений. 40, 604–612. doi: 10.1093/oxfordjournals.pcp.a029583
CrossRef Full Text | Google Scholar
Gajdanowicz, P., Michard, E., Sandmann, M., Rocha, M., Corrêa, L.G., Ramirez-Aguilar, S. J., et al. (2011). Градиенты калия (К+) служат мобильным источником энергии в сосудистых тканях растений. Проц. Натл. акад. науч. США 108, 864–869. doi: 10.1073/pnas.1009777108
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Гальвани, Л. (1791 г.). De viribus electricitatis в комментариях к Motu Muscleis. Bononiensi Sci. Артиум инст. атк акад. Комментарий. 7, 363–418. doi: 10.1097/00000441-195402000-00023
CrossRef Full Text
Geiger, D., Becker, D., Vosloh, D., Gambale, F., Palme, K., Rehers, M., et al. (2009). Гетеромерные каналы AtKC1 {средняя точка} AKT1 в корнях арабидопсиса облегчают рост в условиях ограничения K +. Дж. Биол. Химия . 284, 21288–21295. дои: 10.1074/jbc.M109.017574
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Гербер С. Х. и Зюдхоф Т. К. (2002). Молекулярные детерминанты регулируемого экзоцитоза. Диабет 51, 3–11. doi: 10.2337/diabetes.51.2007. S3
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Гиль, Р. Ф., и фон Вирен, Н. (2014). Корневая подкормка питательными веществами. Завод физиол. 166, 509–517. doi: 10.1104/pp.114.245225
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Гласс, А.Д., Шафф, Дж.Е., и Кочиан, Л.В. (1992). Исследования поглощения нитратов ячменем: IV. Электрофизиология. Завод физиол. 99, 456–463.
Реферат PubMed | Google Scholar
Гоберт А., Исаенков С., Фелькер К., Чемпински К. и Маатуис Ф. Дж. М. (2007). Ген двухпорового канала TPK1 кодирует вакуолярную проводимость K + и играет роль в гомеостазе K + . Проц. Натл. акад. науч. США . 104, 10726–10731. doi: 10.1073/pnas.0702595104
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Goldsworthy, A. (1983). Эволюция потенциалов действия растений. Дж. Теор. биол. 103, 645–648.
Грэмс Т. Е., Лотнер С., Фелле Х. Х., Матиссек Р. и Фромм Дж. (2009). Электрические сигналы, индуцированные теплом, влияют на pH цитоплазмы и апопласта, а также на фотосинтез при распространении через лист кукурузы. Окружающая среда растительных клеток. 32, 319–326. doi: 10.1111/j.1365-3040.2008.01922.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Грубер Б.Д., Гиль Р.Ф.Х., Фридель С. и фон Вирен Н. (2013). Пластичность корневой системы арабидопсиса при дефиците питательных веществ. Завод физиол. 163, 161–179. doi: 10.1104/pp.113.218453
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Гунсе Б., Пошенридер К., Ранкл С., Шредер П., Родриго-Морено А. и Барсело Дж. (2016). Универсальная и легко настраиваемая система для электрофизиологии растений. Методы 3, 436–451. doi: 10.1016/j.mex.2016.05.007
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Guo, J. , Zeng, W., Chen, Q., Lee, C., Chen, L., Yang, Y., et al. (2016). Структура потенциалзависимого двухпорового канала TPC1 из Arabidopsis thaliana . Природа 531, 196–201. doi: 10.1038/nature16446
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Hedrich, R. (2012). Ионные каналы в растениях. Физиол. Откр. 92, 1777–1811 гг. doi: 10.1152/physrev.00038.2011
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Хедрих Р., Сальвадор-Рекатала В. и Дрейер И. (2016). Электропроводка и междугородная связь завода. Trends Plant Sci. 21, 376–387. doi: 10.1016/j.tplants.2016.01.016
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Hegenauer, V., Fürst, U., Kaiser, B., Smoker, M., Zipfel, C., Felix, G., et al. (2016). Обнаружение паразита растений Cuscuta reflexa с помощью рецептора на поверхности клеток томата. Наука 353, 478–481. doi: 10.1126/science. aaf3919
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Хо, С.-Х., Лин, С.-Х., Ху, Х.-С., и Цай, Ю.-Ф. (2009). CHL1 функционирует как датчик нитратов в растениях. Сотовый 138, 1184–1194. doi: 10.1016/j.cell.2009.07.004
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ходжкин, А.Л. (1937). Доказательства электрической передачи в нерве. Журнал физиол. 90, 183–210.
Google Scholar
Ходжкин А.Л. и Хаксли А.Ф. (1952). Количественное описание мембранного тока и его приложение к проводимости и возбуждению в нерве. J. Physiol. 117, 500–544.
Реферат PubMed | Google Scholar
Хоуп, А. Б. (1961a). Ионные отношения клеток chara australis. австр. Дж. Биол. науч. 14, 312–322. doi: 10.1071/BI9610312
Полный текст CrossRef | Академия Google
Хоуп, А.Б. (1961b). Потенциал действия в клетках хара. Природа 191, 811–812. doi: 10.1038/191811a0
Полный текст CrossRef | Google Scholar
Хоуп, А. Б., и Финдли, Г. П. (1964). Потенциал действия у Чары . Физиология клеток растений. 5, 377–380. doi: 10.1093/oxfordjournals.pcp.a079056
CrossRef Full Text | Google Scholar
Hosy, E., Vavasseur, A., Mouline, K., Dreyer, I., Gaymard, F., Porée, F., et al. (2003). Arabidopsis наружный K + канал GORK участвует в регуляции устьичных движений и транспирации растений. Проц. Натл. акад. науч. США . 100, 5549–5554. doi: 10.1073/pnas.0733970100
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Джексон, МБ (2006). Молекулярная и клеточная биофизика . Кембридж: Издательство Кембриджского университета.
Google Scholar
Джулиус Д. и Натанс Дж. (2012). Передача сигналов сенсорными рецепторами. Гавань Колд Спринг. Перспектива. биол. 4:а005991. doi: 10.1101/cshperspect.a005991
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Канчисвами, К. Н., Малной, М., Окчипинти, А., и Маффей, М. Э. (2014). Перспективы визуализации кальция в растениях. Междунар. Дж. Мол. науч. 15, 3842–3859. doi: 10.3390/ijms15033842
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кандель, Э. Р. (2001). Молекулярная биология хранения памяти: диалог между геном и синапсами. Наука 294, 1030–1038. doi: 10.1126/science.1067020
Полный текст CrossRef | Google Scholar
Карбан, Р. (2015). Датчики растений и связь . Чикаго, Иллинойс: University of Chicago Press.
Google Scholar
Катерия С., Нагель Г., Бамберг Э. и Хегеманн П. (2004). «Зрение» у одноклеточных водорослей. Новости Физиол. науч. 19, 133–137. doi: 10.1152/nips.01517.2004
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Киба Т., Кудо Т., Кодзима М. и Сакакибара Х. (2011). Гормональный контроль усвоения азота: роль ауксина, абсцизовой кислоты и цитокинина. Дж. Экспл. Бот. 62, 1399–1409. doi: 10.1093/jxb/erq410
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ким С.А., Квак Дж.М., Джэ С.К., Ван М.Х. и Нам Х.Г. (2001). Сверхэкспрессия гена AtGluR2 , кодирующего гомолог арабидопсиса глутаматных рецепторов млекопитающих, нарушает утилизацию кальция и чувствительность к ионному стрессу у трансгенных растений. Физиол клеток растений . 42, 74–84. doi: 10.1093/pcp/pce008
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Киркби Э. (2011). «Введение, определение и классификация питательных веществ», в Marschner’s Mineral Nutrition of Higher Plants, 3rd Edn ., ed P. Marschner (London: Elsevier), 3–5. doi: 10.1016/B978-0-12-384905-2.00001-7
Полный текст CrossRef | Google Scholar
Китагава М. и Джексон Д. (2017). Опосредованная плазмодесмами межклеточная коммуникация в апикальной меристеме побега: как разговаривают стволовые клетки. Растения 6:12. doi: 10.3390/plants6010012
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Котула Дж. В., Кернс С. Дж., Шакет Л. А., Сирадж Л., Коллинз Дж. Дж., Уэй Дж. К. и др. (2014). Программируемые бактерии обнаруживают и записывают сигналы окружающей среды в кишечнике млекопитающих. Проц. Натл. акад. науч. США 111, 4838–4843. doi: 10.1073/pnas.1321321111
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Курусу Т., Хамада Х., Кояно Т. и Кучицу К. (2012). Внутриклеточная локализация и физиологическая функция риса Ca 2+ -проницаемый канал OsTPC1. Сигнал завода. Поведение 7, 1428–1430. doi: 10.4161/psb.22086
Полный текст CrossRef | Google Scholar
Лаанемец К., Брандт Б., Ли Дж., Мерило Э., Ван Ю.-Ф., Кешвани М.М. и др. (2013). Кальций-зависимая и независимая устьичная сигнальная сеть и компенсаторная обратная связь, контролирующая открывание устьиц посредством запуска чувствительности к Ca2+. Завод физиол. 163, 504–513. doi: 10.1104/pp.113.220343
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Ли, Дж. Ю. (2015). Plasmodesmata: сигнальный узел на клеточной границе. Курс. мнение биол. растений 27, 133–140. doi: 10.1016/j.pbi.2015.06.019
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ленг К., Мерсье Р. В., Хуа Б.-Г., Фромм Х. и Берковиц Г. А. (2002). Электрофизиологический анализ клонированных ионных каналов, управляемых циклическими нуклеотидами. Завод Физиол . 128, 400–410. doi: 10.1104/pp.010832
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Ленг, К., Мерсье, Р.В., Яо, В., и Берковиц, Г.А. (1999). Клонирование и первая функциональная характеристика катионного канала растений, управляемого циклическим нуклеотидом. Завод Физиол . 121, 753–761.
Реферат PubMed | Google Scholar
Li, Z.Y., Xu, Z.S., He, G.Y., Yang, G. X., Chen, M., Li, L.C., et al. (2013). Потенциал-зависимый анионный канал 1 (AtVDAC1) негативно регулирует реакцию растений на холод во время прорастания и развития проростков арабидопсиса и взаимодействует с кальциевым сенсором CBL1. Междунар. Дж. Мол. Наука . 14, 701–713. doi: 10.3390/ijms14010701
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Лю К., Ли Л. и Луан С. (2006). Внутриклеточное определение K + SKOR, канала K + шейкерного типа из Arabidopsis. Завод J . 46, 260–268. doi: 10.1111/j.1365-313X.2006.02689.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Liu, K.H., Niu, Y., Konishi, M., Wu, Y., Du, H., Sun Chung, H., et al. (2017). Открытие передачи сигналов нитрат-КФК-НЛП в центральных сетях роста питательных веществ. Природа 545, 311–316. doi: 10.1038/nature22077
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Лопес-Миллан А. Ф., Моралес Ф., Абадиа А. и Абадиа Дж. (2001). Ассоциированные с дефицитом железа изменения состава апопластной жидкости листьев полевых деревьев груши ( Pyrus communis L.). Дж. Экспл. Бот. 52, 1489–1498. doi: 10.1093/jexbot/52.360.1489
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Ma, W., Smigel, A., Walker, R.K., Moeder, W., Yoshioka, K., and Berkowitz, G.A. (2010). Передача сигналов старения листьев: Ca2+-проводящий канал 2 арабидопсиса, управляемый циклическими нуклеотидами, действует через оксид азота, подавляя программирование старения. Завод Физиол . 154, 733–743. doi: 10.1104/pp.110.161356
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Maathuis, FJ (2009). Физиологические функции минеральных макроэлементов. Курс. мнение биол. растений 12, 250–258. doi: 10.1016/j.pbi.2009.04.003
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мальческа Ф. , Ахмад А., Батул С., Мюллер Х. М., Людвиг-Мюллер Дж., Кройцвизер Дж. и др. (2017). Сульфат ксилемного сока, усиленный засухой, закрывает устьица, влияя на ALMT12 и синтез ABA замыкающих клеток. Завод физиол. 174, 798–814. doi: 10.1104/pp.16.01784
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мартинак, Б. (2008). Зондирование с помощью ионных каналов . изд. Б. Мартинак. Берлин; Гейдельберг: Спрингер.
Google Scholar
Маси Э. и Цишак М. (2014). Электрические выбросы в бактериальных биопленках. JR Soc. Интерфейс 12:20141036. doi: 10.1098/rsif.2014.1036
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Masi, E., Ciszak, M., Comparini, D., Monetti, E., Pandolfi, C., Azzarello, E., et al. (2015). Электрическая сеть верхушки корня кукурузы зависит от гравитации. Науч. Респ. 5:7730. doi: 10.1038/srep07730
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
McClure, P. R., Kochian, L.V., Spanswick, R.M., and Shaff, JE (1990). Доказательства котранспорта нитратов и протонов в корнях кукурузы: I. Влияние нитратов на мембранный потенциал. Завод физиол. 93, 281–289.
Реферат PubMed | Google Scholar
Мехарг А.А. и Блатт М.Р. (1995). NO3-транспорт через плазматическую мембрану Корневые волоски Arabidopsis thaliana : кинетический контроль с помощью рН и мембранного напряжения. Дж. Мембр. биол. 145, 49–66. doi: 10.1007/BF00233306
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мерчант С. С., Прочник С. Э., Валлон О., Харрис Э. Х., Карпович С. Дж., Уитман Г. Б. и др. (2007). Геном Chlamydomonas раскрывает эволюцию ключевых функций животных и растений. Наука 318, 245–250. doi: 10.1126/science.1143609
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Мейер С., Мумм П., Имес Д., Эндлер А., Ведер Б., Аль-Рашейд К. А. и др. (2010). AtALMT12 представляет собой анионный канал R-типа, необходимый для движения устьиц в замыкающих клетках арабидопсиса. Завод J . 63, 1054–1062. doi: 10.1111/j.1365-313X.2010.04302.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Морган, А. Дж., и Галионе, А. (2014). Двухпоровые каналы (TPC): текущие споры. Bioessays 36, 173–183. doi: 10.1002/bies.201300118
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Мулин К., Вери А.-А., Гаймар Ф., Бушерез Дж., Пилот Г., Девик М. и др. (2002). Развитие пыльцевых трубок и конкурентоспособность нарушаются из-за разрушения канала Shaker K + в Arabidopsis . Гены Дев . 16, 339–350. doi: 10.1101/gad.213902
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мусави С.А., Шовен А., Паско Ф., Келленбергер С. и Фармер Э.Э. (2013). Гены GLUTAMATE RECEPTOR-LIKE опосредуют передачу сигналов ран от листа к листу. Природа 500, 422–426. doi: 10.1038/nature12478
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мучхал, США, и Раготама, К. Г. (1999). Транскрипционная регуляция переносчиков фосфатов растений. Проц. Натл. акад. науч. США 96, 5868–5872.
Реферат PubMed | Google Scholar
Накагаки Т. (2001). Умное поведение настоящего слизевика в лабиринте. Рез. микробиол. 152, 767–770. дои: 10.1016/S0923-2508(01)01259-1
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Наврат К., Шрайбер Л., Франке Р. Б., Гельднер Н., Рейна-Пинто Дж. Дж. и Кунст Л. (2013). Апопластические диффузионные барьеры у арабидопсиса. араб. Б. 11:e0167. doi: 10.1199/tab.0167
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Окубо Ю., Танака М., Табата Р., Огава-Ониши М. и Мацубаяши Ю. (2017). Мобильные полипептиды от побегов к корням, участвующие в системной регуляции усвоения азота. Нац. Plants 3, 17029. doi: 10.1038/nplants.2017.29
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Peiter, E., Maathuis, FJM, Mills, L. N., Knight, H., Pelloux, J., Hetherington, A.M., et al. (2005). Вакуолярный Ca 2+ -активируемый канал TPC1 регулирует прорастание и движение устьиц. Природа 434, 404–408. doi: 10.1038/nature03381
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Пере, Б., Клеман, М., Нуссоме, Л., и Деснос, Т. (2011). Адаптация развития корней к фосфатному голоданию: лучше перестраховаться, чем потом сожалеть. Тенденции Растениевод. 16, 442–450. doi: 10.1016/j.tplants.2011.05.006
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Пикард, Б.Г. (1973). Потенциалы действия у высших растений. Бот. Ред. 39, 172–201. doi: 10.1007/BF02859299
Полный текст CrossRef | Google Scholar
Петрушка М., Столарек Я. А. Н. и Пазуркевич-Кокот К. (1997). Эволюция потенциала действия в растительных клетках во времени. Дж. Биол. физ. 23, 219–232.
Реферат PubMed | Google Scholar
Приндл А. , Лю Дж., Асалли М., Ли С., Гарсия-Охальво Дж., Зуэль Г. М. и др. (2015). Ионные каналы обеспечивают электрическую связь в бактериальных сообществах. Природа 527, 59–63. doi: 10.1038/nature15709
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Риверас Э., Альварес Дж. М., Видаль Э. А., Осес К., Вега А. и Гутьеррес Р. А. (2015). Ион кальция является вторичным мессенджером в сигнальном пути нитратов арабидопсиса. Физиол растений. 169, 1397–1404. doi: 10.1104/pp.15.00961
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Rocchetti, A., Sharma, T., Wulfetange, C., Scholz-Starke, J., Grippa, A., Carpaneto, A., et al. (2012). Предполагаемая субъединица канала K(+) AtKCO3 образует стабильные димеры у арабидопсиса. Передний завод Sci . 3:251. doi: 10.3389/fpls.2012.00251
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ронзье Э., Корратже-Файе К., Санчес Ф., Прадо К. , Бриер К., Леонхардт Н. и др. (2014). CPK13, неканоническая Ca2+-зависимая протеинкиназа, специфически ингибирует каналы KAT2 и KAT1shaker K+ и уменьшает открывание устьиц. Завод Физиол . 166, 314–326. doi: 10.1104/pp.114.240226
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Рубио В., Линьярес Ф., Солано Р., Мартин А. К., Иглесиас Дж., Лейва А. и др. (2001). Консервативный фактор транскрипции MYB, участвующий в передаче сигналов фосфатного голодания как у сосудистых растений, так и у одноклеточных водорослей. Гены Дев. 15, 2122–2133. doi: 10.1101/gad.204401
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Сакано, К. (1990). Протонно-фосфатная стехиометрия поглощения неорганического фосфата культивируемыми клетками Catharanthus roseus (L.) G. Don. Завод физиол. 93, 479–83.
Реферат PubMed | Google Scholar
Сальвадор-Рекатала, В. (2016a). Новые роли генов, подобных глутаматным рецепторам 3. 3, 3.5 и 3.6, в качестве переключателей системных электрических сигналов, вызванных раной. Сигнал завода. Поведение 11:e1161879. дои: 10.1080/15592324.2016.1161879
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Сальвадор-Рекатала, В. (2016b). Калиевый канал AKT2 опосредует индуцированную NaCl деполяризацию в корне Arabidopsis thaliana . Сигнал завода. Поведение 11:e1165381. doi: 10.1080/15592324.2016.1165381
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Сальвадор-Рекатала, В., Тьяллинги, В. Ф., и Фармер, Э. Э. (2014). В реальном времени, in vivo внутриклеточные записи индуцированных гусеницами волн деполяризации в ситовидных элементах с использованием тлей-электродов. Новый фитол. 203, 674–684. doi: 10.1111/nph.12807
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Сандерсон, Дж. Б. (1872 г.). Примечание об электрических явлениях, сопровождающих раздражение листа Dionaea muscipula . Проц. R Соц. Лонд. 21, 495–496. doi: 10.1098/rspl.1872.0092
CrossRef Full Text
Sattelmacher, B. (2001). Апопласт и его значение для минерального питания растений. Новый фитол. 149, 167–192. doi: 10.1046/j.1469-8137.2001.00034.x
Полный текст CrossRef | Google Scholar
Саттельмахер Б. и Хорст В. Дж. (2007). Апопласт высших растений: отделение хранения, транспорта и реакций: значение апопласта для минерального питания высших растений. Дордрехт: Спрингер.
Google Scholar
Шахтман Д. П. и Шин Р. (2007). Определение питательных веществ и сигнализация: NPKS. год. Преподобный завод биол. 58, 47–69. doi: 10.1146/annurev.arplant.58.032806.103750
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Шредер Дж. И., Хедрих Р. и Фернандес Дж. М. (1984). Калий-селективные одиночные каналы в протопластах замыкающих клеток Vicia faba . Природа 312, 361–362. doi: 10.1038/312361a0
CrossRef Полный текст | Google Scholar
Шахзад З. и Амтманн А. (2017). Пища для размышлений: как питательные вещества регулируют архитектуру корневой системы. Курс. мнение биол. растений 39, 80–87. doi: 10.1016/j.pbi.2017.06.008
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Сомссич М., Хан Г. А. и Перссон С. (2016). Неоднородность клеточной стенки в развитии корней арабидопсиса. Фронт. Растениевод. 7:1242. doi: 10.3389/fpls.2016.01242
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Спэнсвик, Р. М., и Костертон, Дж. В. Ф. (1967). Плазмодесмы в Nitella translucens : структура и электрическое сопротивление. J. Cell Sci. 2, 451–464.
Реферат PubMed | Google Scholar
Страль, Х., и Хамоен, Л.В. (2010). Мембранный потенциал важен для деления бактериальных клеток. Проц. Натл. акад. науч. США . 107, 12281–12286. doi: 10.1073/pnas.1005485107
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Широкий А. , Ивашикина Н., Дитрих П., Рулфсема М. Р. Г., Аче П., Рейнтанц Б. и др. (2001). KAT1 не важен для открывания устьиц. Проц. Натл. акад. науч. США . 98, 2917–2921. doi: 10.1073/pnas.051616698
CrossRef Full Text | Google Scholar
Татеда К., Ватанабэ К., Кусано Т. и Такахаши Ю. (2011). Молекулярно-генетическая характеристика семейства генов, кодирующих потенциалзависимый анионный канал у арабидопсиса. Дж. Экспл. Бот . 62, 4773–4785. doi: 10.1093/jxb/err113
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Тейлор, А. Р., Браунли, К., и Уиллер, Г. Л. (2012). Протонные каналы в водорослях: поводы для волнения. Тенденции Растениевод. 17, 675–684. doi: 10.1016/j.tplants.2012.06.009
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Тилснер Дж., Николя В., Росадо А. и Байер Э. М. (2016). Оставаться на месте: места контакта с плазмодесмальной мембраной и контроль связи между клетками у растений. год. Преподобный завод биол. 67, 1–28. doi: 10.1146/annurev-arplant-043015-111840
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Требач К. и Завадски Т. (1985). Световые потенциалы действия у печеночника Conocephalum conicum . Физиол. Завод. 64, 482–486. doi: 10.1111/j.1399-3054.1985.tb08526.x
Полный текст CrossRef | Google Scholar
Ульрих-Эбериус К.И., Новацкий А., Фишер Э. и Люттге У. (1981). Взаимосвязь между энергозависимым поглощением фосфатов и электрическим потенциалом мембраны ряски гибба G1. Завод физиол. 67, 797–801.
Реферат PubMed | Академия Google
Видаль, Э.А., Араус, В., Лу, К., Парри, Г., Грин, П.Дж., Коруцци, Г.М., и соавт. (2010). Регуляторный модуль miR393/AFB3, чувствительный к нитратам, контролирует архитектуру корневой системы у Arabidopsis thaliana . Проц. Натл. акад. науч. США 107, 4477–4482. doi: 10.1073/pnas.01107
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Фелькер К. , Шмидт Д., Мюллер-Робер Б. и Чемпински К. (2006). Члены семейства AtTPK/KCO арабидопсиса образуют гомомерные вакуолярные каналы на заводе . Завод J . 48, 296–306. doi: 10.1111/j.1365-313X.2006.02868.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Волков А.Г. (2012). Электрофизиология растений . ред. А. Г. Волкова. Берлин; Гейдельберг: Спрингер.
Реферат PubMed | Google Scholar
фон Вангенхайм Д., Го Т., Дитрих Д. и Беннет М. Дж. (2017). Биология растений: создание барьеров… в корнях. Курс. биол. 27, Р172–Р174. doi: 10.1016/j.cub.2017.01.060
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Waese, J., Fan, J., Pasha, A., Yu, H., Fucile, G., Shi, R., et al. (2017). ePlant: визуализация и изучение нескольких уровней данных для генерации гипотез в биологии растений. Растительная клетка 29, 1806–1821. doi: 10.1105/tpc.17.00073
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ван Р. и Кроуфорд Н. М. (1996). Генетическая идентификация гена, участвующего в конститутивном высокоаффинном транспорте нитратов у высших растений. Проц. Натл. акад. науч. США 93, 9297–9301.
Реферат PubMed | Google Scholar
Ван Р., Лю Д. и Кроуфорд Н. М. (1998). Белок CHL1 арабидопсиса играет важную роль в высокоаффинном поглощении нитратов. Проц. Натл. акад. науч. США 95, 15134–15139.
Реферат PubMed | Google Scholar
Ван, Р., Окамото, М., Син, X., и Кроуфорд, Н. М. (2003). Микроматричный анализ реакции нитратов корней и побегов арабидопсиса выявил более 1000 быстро реагирующих генов и новые связи с метаболизмом глюкозы, трегалозо-6-фосфата, железа и сульфатов. Физиол растений. 132, 556–567. doi: 10.1104/pp.103.021253
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Уорд, Дж. М., Мезер, П., и Шредер, Дж. И. (2009). Ионные каналы растений: семейства генов, физиология и анализ функциональной геномики. год. Преподобный Физиол. 71, 59–82. doi: 10.1146/annurev.physiol.010908.163204
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Уилер Г.Л. и Браунли К. (2008). Передача сигналов Ca2+ в растениях и зеленых водорослях — изменение каналов. Тенденции Растениевод. 13, 506–514. doi: 10.1016/j.tplants.2008.06.004
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Xu, J., Li, H.-D., Chen, L.-Q., Wang, Y., Liu, L.-L., He, L., et al. (2006). Протеинкиназа, взаимодействуя с двумя кальцинейрин В-подобными белками, регулирует транспортер K + AKT1 в Arabidopsis . Сотовый 125, 1347–1360. doi: 10.1016/j.cell.2006.06.011
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Ян Дж., Хе Х., Тонг С., Чжан В., Ван Дж., Ли Х. и др. (2009). Потенциал-зависимый анионный канал 2 Arabidopsis thaliana (AtVDAC2) участвует в опосредованном АБК раннем развитии проростков. Междунар. Дж. Мол. Наука . 10, 2476–2486. doi: 10.3390/ijms10062476
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ян В., Нагасава К., Мюнч К., Сюй Ю., Саттерстром К., Чон С. и др. (2016). Сеть митохондриальных сиртуинов обнаруживает динамическое SIRT3-зависимое деацетилирование в ответ на деполяризацию мембраны. Сотовый 0, 14447–14452. doi: 10.1016/j.cell.2016.10.016
CrossRef Полный текст | Google Scholar
Yang, X. Y., Chen, Z. W., Xu, T., Qu, Z., Pan, X. D., Qin, X. H., et al. (2011). Кинезин КР1 арабидопсиса специфически взаимодействует с митохондриальным белком VDAC3 и регулирует дыхание при прорастании семян при низкой температуре. Растительная клетка 23, 1093–1106. doi: 10.1105/tpc.110.082420
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Завадски Т., Дэвис Э., Бзюбинска Х., Хткбач К. и Характеристики К. (1991). Характеристики потенциала действия Helianthus annuus . Физиол. Завод. 83, 601–604.
Google Scholar
Зебело, С. А., Мацуи, К., Одзава, Р., и Маффей, М. Э. (2012). Деполяризация потенциала плазматической мембраны и цитозольный поток кальция являются ранними событиями, участвующими в коммуникации томата ( Solanum lycopersicon ) между растениями. Растениевод. 196, 93–100. doi: 10.1016/j.plantsci.2012.08.006
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чжан А., Жэнь Х.-М., Тан Ю.-К., Ци Г.-Н., Яо Ф.-Ю., Ву Г.-Л. и др. др. (2016). Анионные каналы S-типа SLAC1 и SLAH4 функционируют как важные негативные регуляторы внутренних каналов K + и открытия устьиц у Arabidopsis . Растительная клетка 28, 949–965. doi: 10.1105/tpc.16.01050
CrossRef Full Text | Академия Google
Чжан Дж., Мартинойя Э. и Де Анджели А. (2014). Цитозольные нуклеотиды блокируют и регулируют вакуолярный анионный канал арабидопсиса AtALMT9. Дж. Биол. хим. 289, 25581–25589. doi: 10.1074/jbc.M114.576108
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чжан X. и Чжан Ю. (2009). Нервно-иммунная коммуникация у Caenorhabditis elegans. Микроб-хозяин клетки 5, 425–429. doi: 10.1016/j.chom.2009.05.003
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Чжэн, X., Хэ, К., Клейст, Т., Чен, Ф., и Луань, С. (2015). Анионный канал SLAh4 участвует в нитрат-зависимом снижении токсичности аммония у арабидопсиса. Окружающая среда растительных клеток . 38, 474–486. doi: 10.1111/pce.12389
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чжэн, З.-Л. (2009). Сигнализация баланса питательных веществ углерода и азота в растениях. Сигнал завода. Поведение 4, 584–591. doi: 10.4161/psb.4.7.8540
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Чжоу В., Брокмеллер Т., Линг З., Омдал А., Болдуин И. Т. и Сюй С. (2016). Эволюция передачи сигналов ранней защиты, индуцированной травоядными, была сформирована полногеномными дупликациями у Nicotiana. eLife 5:e19531. doi: 10.7554/eLife.19531
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Циммерманн М. Р., Майшак Х., Митхофер А., Боланд В. и Фелле Х. Х. (2009). Системные потенциалы, новый электрический апопластический сигнал на большие расстояния у растений, вызванный ранением. Физиол растений. 149, 1593–1600. doi: 10.1104/pp.108.133884
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Новый способ запечатлеть электрическую симфонию мозга
Биофизик Адам Коэн прогуливался по Сан-Франциско, штат Калифорния, в 2010 году, когда телефонный звонок застал его врасплох. «У нас есть сигнал», — сказал звонивший. Почти в 5000 километрах от него, в Кембридже, штат Массачусетс, его сотрудники нашли золото. После нескольких месяцев неудачных экспериментов исследователи нашли флуоресцентный белок, который позволил им наблюдать сигналы, проходящие между нейронами.
Но происходило что-то странное. Когда Коэн вернулся в свою лабораторию в Гарвардском университете, он узнал, что все записи эксперимента демонстрируют странную прогрессию. Сначала нейроны, украшенные белком, красиво вспыхивали, когда через них проносились электрические импульсы. Но затем клетки превратились в яркие пятна. «На полпути к каждой записи сигнал становился диким», — говорит Коэн.
Поэтому он решил присоединиться к своей команде во время эксперимента. «Когда они начинали запись, они сидели, затаив дыхание», — говорит Коэн. Но как только они понимали, что это работает, они праздновали, «танцевая и бегая по комнате».
В своем изобилии они пускали свет настольной лампы прямо в микроскоп. «На самом деле мы записывали наше волнение», — говорит Дэниел Хохбаум, в то время аспирант группы Коэна. Они смягчили свои празднования, и год спустя команда опубликовала свое исследование 1 — одно из первых, показавшее, что флуоресцентный белок, встроенный в определенные нейроны млекопитающих, можно использовать для отслеживания отдельных электрических импульсов в реальном времени.
Нейробиологи десятилетиями пытались наблюдать за быстрыми электрическими сигналами, которые являются основным компонентом языка мозга. Хотя электроды, рабочая лошадка для измерения напряжения, могут надежно регистрировать активность отдельных нейронов, они с трудом улавливают сигналы многих, особенно в течение длительных периодов времени. Но за последние два десятилетия ученые нашли способ встраивать флуоресцентные белки, указывающие напряжение, прямо в клеточные мембраны нейронов. С помощью подходящего микроскопа они смогут увидеть, как клетки загораются, когда они разговаривают друг с другом — будь то шепот или крик. Визуализация напряжения также может регистрировать электрические колебания между многими нейронами одновременно, а затем усреднять эти сигналы по большим участкам мозговой ткани. Это помогает исследователям изучать электрическую активность мозга в разных пространственных масштабах, слушая не только голоса отдельных клеток, но и «рев толпы», говорит Коэн.
За последние 5 лет ученые опубликовали около 1000 статей по этой теме, а крупные схемы финансирования, такие как инициатива BRAIN Национального института здравоохранения США, ускорили разработку новых типов генно-инженерных индикаторов напряжения. В надежде найти лучшие варианты некоторые группы придумали стратегии для скрининга миллионов белков на наличие желаемых характеристик, таких как яркость. Один из таких подходов определил индикатор, который в два раза ярче, чем аналогичные датчики, разработанные всего четырьмя годами ранее 9.0035 2 .
По мере того, как эти белки улучшаются, а достижения в микроскопии облегчают их наблюдение, ученые надеются пролить свет на самую большую загадку нейронауки: как клетки мозга работают вместе, чтобы преобразовать систему электрических импульсов в мысли, действия и эмоции. Исследователи все еще пытаются уловить весь спектр активности и разработать способы увидеть быстрое и глубокое возбуждение нервов в тканях мозга. Но если достижения смогут решить эти технические проблемы, «это будет революционно», — говорит Рафаэль Юсте, изучающий функцию нейронных цепей в Колумбийском университете в Нью-Йорке.
Высокоскоростной процесс
Средний человеческий мозг содержит около 120 миллиардов нейронов, которые постоянно получают и отправляют информацию через разветвленные придатки, называемые дендритами. Химические или электрические сигналы, которые достигают дендритов, вызывают небольшие изменения напряжения на клеточной мембране, которые направляются к телу клетки. Когда сумма изменений напряжения достигает точки невозврата, называемой порогом, нейрон запускает большой электрический импульс — потенциал действия. Этот толчок распространяется со скоростью до 150 метров в секунду вдоль нейронной ветви, известной как аксон, к другому набору ветвящихся придатков. Здесь химические или электрические сигналы передают информацию следующему набору дендритов.
100 летучих мышей и длинный темный туннель: попытка одного нейробиолога раскрыть секреты трехмерной навигации
Нейронные сигналы сходятся, расходятся и синхронизируются, создавая симфонию мыслей, эмоций, действий и реакций, от покраснения лица до детской икоты. Но инструменты прослушивания ученых крайне ограничены. Миниатюрные электроды толщиной с волос, впервые разработанные в 1940-х годах, можно вводить в мозг, напротив или внутри нейронов, где они измеряют мембранное напряжение с точностью и скоростью. Но этот подход можно использовать для мониторинга только одного или нескольких нейронов одновременно — и только в течение ограниченного периода времени, потому что электроды в конечном итоге повреждают клетку. Это все равно, что пытаться уловить суть оркестровой аранжировки, следя за одним исполнителем в течение нескольких секунд.
Связки микроэлектродов могут регистрировать электрическую активность до 200 клеток одновременно, но поскольку эти электроды расположены рядом с нейронами, а не внутри них, они могут регистрировать только потенциалы действия, самые резкие всплески электрической активности. Они глухи к более мягким нотам — небольшим электрическим изменениям, которые не подталкивают нейрон к потенциалу действия. Эти подпороговые изменения напряжения являются ключевыми для работы мозга, потому что они постепенно складываются, чтобы определить, сработает нейрон или нет.
В надежде измерить более тихую мозговую активность в больших популяциях клеток ученые в 1960-х годах начали экспериментировать с идеей сенсора или зонда, который флуоресцирует в ответ на электрический сигнал. Самые популярные зонды, называемые кальциевыми индикаторами, загораются при связывании с кальцием, поступающим в нейрон в результате всплеска электрической активности. Но этот метод, известный как кальциевая визуализация, дает только косвенное представление; он не записывает непосредственно мембранное напряжение. И хотя он покажет сигнал крупных событий, таких как потенциалы действия, он упускает важные для работы мозга вещи, такие как тонкие колебания мембранного напряжения или электрические сигналы, подавляющие потенциалы действия. Представьте себе, что после симфонического концерта можно услышать только взрыв аплодисментов: ясно, что оркестр выступал, но что он играл, остается только гадать.
В 1970-х годах ученые начали разрабатывать сенсоры на красителях, которые напрямую обнаруживают изменения в мембранном напряжении. Первые версии этих красителей должны были быть нанесены на мозг без разбора, поэтому они помечали все типы клеток, включая ненейронные клетки, что затрудняло анализ активности конкретных нейронов.
Затем, в 1990-х годах, исследователи начали тестировать индикаторы, которые можно было сконструировать с помощью генной инженерии, чтобы они проявлялись только в интересующих нейронах. Первый 3 генетически кодируемый индикатор напряжения (GEVI) был разработан в 1997; с тех пор ученые выпустили более двух десятков датчиков 4 . Некоторые из них производятся путем объединения белка, чувствительного к напряжению, с флуоресцентными молекулами (см. «Ароматы флуоресценции»). Когда эти белки обнаруживают изменение напряжения, они меняют свою трехмерную структуру и изменяют флуоресценцию молекулы, с которой они связаны. Другими индикаторами напряжения являются мутированные версии микробных родопсинов, флуоресцентных молекул, которые вызывают изменение напряжения на плазматической мембране в ответ на свет. Эти белки также могут работать в обратном направлении, изменяя свою реакцию на свет — и, следовательно, свою флуоресценцию — в ответ на изменение напряжения мембраны.
Все в деталях
На данный момент GEVI доказали свою эффективность в отслеживании индивидуальных потенциалов действия как в культивируемых нейронах, выращенных в чашке, так и в интактном мозге широкого круга животных, от насекомых 5 до мышей 6 . По словам Коэна, одним из самых больших обещаний метода является его способность регистрировать не только крупные события, но и небольшие подпороговые изменения напряжения мембраны, которые отражают сообщения, которые нейрон получает от соседних клеток. «Визуализация напряжения позволяет увидеть входные данные нейронов in vivo , на который раньше у нас не было возможности взглянуть», — говорит он.
В прошлом году Коэн и его коллеги разработали новые GEVI и улучшили методы микроскопии для регистрации таких подпороговых изменений напряжения от многих нейронов одновременно, в том числе в мозге мыши 7 , 8 . Команда также смогла записать электрическую активность тех же нейронов через неделю. Способность точно знать, какие нейроны записываются, и отслеживать их во времени позволяет исследователям смотреть на проводку между этими нейронами, говорит Эд Бойден, нейробиолог из Массачусетского технологического института в Кембридже. Поступая таким образом, «вы можете связать структуру мозга с его функциями», — говорит он. «Это один из основных вопросов во всей неврологии».
Еще одним преимуществом GEVI является то, что, в отличие от электродов, которые регистрируют в основном сигналы от тела клетки, они могут регистрировать электрические сигналы от любой части нервной клетки, вплоть до кончиков дендритов (см. «Попадание на весы»). Это все равно, что слушать ноты, которые играет левая рука пианиста. «Это то, о чем я давно мечтал — и я не одинок», — говорит Каталин Тот, нейробиолог из Университета Лаваля в Квебеке, Канада. По ее словам, многие нейробиологи стремятся отслеживать напряжение во всех нейронах, чтобы увидеть, как оно меняется в разных областях клетки.
Вей Вей, нейробиолог из Чикагского университета, штат Иллинойс, использует GEVI, чтобы выяснить, как различные электрические входы интегрируются в нейроны сетчатки мыши. Вей интересуется классом нейронов, которые сильнее реагируют на зрительный стимул, когда он движется в определенном направлении. Глядя на то, как меняется напряжение мембраны в разных частях этих нейронов, она надеется понять, как клетки суммируют входящие сигналы, чтобы определить направление движения.
Нейрофизиолог Винсент Виллетт из Высшей нормальной школы в Париже планирует использовать датчики напряжения для изучения того, как регулярные колебания подпороговых электрических сигналов определяют, как нейроны в мозжечке мыши координируют мышечную активность. «Многое предстоит понять о том, как клетки действуют вместе», — говорит Виллетт.
Визуальное считывание напряжения мембраны также позволяет ученым видеть электрические сигналы, которые подавляют активацию нейронов, а не запускают ее. Поскольку тормозные сигналы невозможно зарегистрировать с помощью таких подходов, как визуализация кальция, неясно, как именно они формируют активность мозга, говорит Роза Коссар, нейробиолог из Средиземноморского института нейробиологии в Марселе, Франция.
Коссар уже много лет использует электроды и кальциевую визуализацию, но теперь ей не терпится попробовать GEVI. Она надеется, что эти датчики позволят ей измерять напряжение с высокой скоростью на нескольких нейронах — по крайней мере, 50 — одновременно у живой мыши. По ее словам, это помогло бы понять, как группы нейронов интегрируют электрические сигналы — как возбуждающие, так и тормозные — для поддержки деятельности, имеющей решающее значение для развития и функционирования мозга.
Глубокие вызовы
Несмотря на большие ожидания, заставить GEVI работать в лаборатории может быть непросто. Возьмем, к примеру, Хелен Янг: будучи аспиранткой Калифорнийского Стэнфордского университета, она решила попробовать GEVI как способ изучения нейронов зрительной системы плодовой мушки. Но глядя в микроскоп во время своего первого эксперимента, Ян не увидела изменений во флуоресценции клеток, даже когда она посветила ярким светом в глаза мух. Только когда она проанализировала данные, она поняла, что визуальные стимулы генерировали сигнал, он был просто крошечным. «Я была очень взволнована, но мои коллеги по лаборатории были в меньшей степени, — говорит она. «Ответы были довольно маленькими и шумными».
Ян начал играть с настройками микроскопа, увеличивая мощность лазера и ускоряя получение изображений. «По сути, я заставила его работать так быстро, как только мог наш микроскоп», — говорит она. Это потому, что реакция индикатора на электрический сигнал была настолько быстрой, что изменение флуоресценции можно было обнаружить всего за долю секунды. «Если вы снимаете только один кадр в то время, когда ячейка отвечает, ответ совсем не выглядит большим», — говорит Ян.
Как мигающие огни и розовый шум могут избавить от болезни Альцгеймера, улучшить память и многое другое
В конце концов Ян удалось использовать GEVI для изучения того, как нейроны мух обрабатывают визуальные сигналы 5 , но те проблемы, с которыми она столкнулась, до сих пор не позволили визуализации напряжения стать основным методом. По словам Коэна, для этого требуются передовые, часто изготавливаемые на заказ платформы для микроскопов. «Вы не можете просто сделать это на флуоресцентном микроскопе вашей бабушки».
За последние пять лет финансовая поддержка инициативы BRAIN способствовала прогрессу в этой области, включая разработку лучших GEVI, говорит Майкл Лин, инженер-протеин из Стэнфорда.
Параллельно с разработкой новых сенсоров ученые работают над методами точного отображения быстрых электрических сигналов, проходящих через мозг. Одна из проблем заключается в том, что большинство доступных методов хорошо работают только с клетками в чашке или на поверхности мозга. Но мозг млекопитающих не прозрачен: на самом деле он похож на тофу, говорит На Джи, физик из Калифорнийского университета в Беркли.
Чтобы заглянуть глубже, исследователям приходится прибегать к более инвазивным методам, таким как удаление части вышележащей ткани или введение крошечных оптических устройств, называемых микроэндоскопами, непосредственно в мозг. Альтернативным неинвазивным способом исследования непрозрачных тканей — до 1 миллиметра в глубину — является двухфотонная микроскопия. В этом методе используется длинноволновый свет с меньшей энергией, который может проникать глубже в ткани. Поскольку двухфотонные микроскопы освещают и записывают только из одной точки за раз, они захватывают изображения слишком медленно, чтобы отследить большую часть быстрой болтовни мозга. Но специалисты уверены, что развитие технологий вскоре позволит увидеть сигналы, производимые GEVI, на более высокой скорости. «Это абсолютно выполнимо», — говорит Джи.
Если различные подходы смогут решить эти проблемы, ученые не сомневаются, что визуализация напряжения станет стандартным подходом к измерению мозговой активности. «В ближайшие год или два мы увидим много статей, в которых применялись датчики напряжения и изучалась биология», — говорит Томас Клэндинин, нейробиолог из Стэнфорда. Некоторые говорят, что эта техника может даже заменить электроды в вопросах, связанных с тем, как нейроны обрабатывают и интегрируют информацию.
Начинающие исследователи настроены особенно оптимистично: Хохбаум, который в настоящее время является научным сотрудником Гарвардской медицинской школы в Бостоне, говорит, что в долгосрочной перспективе GEVI станет инструментом для изучения того, как различные компартменты клетки реагируют на подпороговые сигналы. Он планирует использовать визуализацию напряжения, чтобы понять, как такие сигналы изменяют связь между нейронами, что является ключевым процессом в обучении. Возможности захватывающие, говорит Хохбаум, но он усвоил по крайней мере один важный урок из тех первых дней, когда он радостно прыгал по лаборатории, увидев свечение в микроскопе: когда эксперименты работают, сведите празднование к минимуму.
Сигналы в электронике – аналоговые и цифровые типы сигналов
Содержание
Электрические сигналы (в электронике) – различные напряжения и токи в электрической сети, называемые электрической «цепью» или «устройством», которые могут быть далее описывается как процесс изменения определенной физической величины или состояния физического объекта в течение определенного периода времени. Используются с целью визуализации, регистрации и передачи сообщений (информации). Сигнал может быть носителем различной информации, например. электрические, магнитные и акустические сигналы и содержит информационный параметр, например. амплитуда, частота или ширина импульса. В электронике наиболее важными сигналами являются изменения электрического заряда, тока, напряжения и электромагнитного поля. Они используются для анализа поведения электронных схем или для измерения изменяющихся электрических величин.
Периодический сигнал — это сигнал, который повторяется в одних и тех же неизменных интервалах времени. Эти интервалы называются периодами сигнала с символом «Т».
Рис. 1. Пример периодической формы сигнала Сигналы в электронике – ПодразделениеСамым основным разделением электрических сигналов является разделение на аналоговые и цифровые сигналы. В электронике различают несколько типов сигналов, к которым относятся:
- Аналоговые сигналы – непрерывные по значениям и времени,
- Цифровые сигналы – дискретные по значениям и времени,
- Дискретные сигналы – непрерывные по значениям, дискретные по времени,
- Квантованные сигналы – дискретные по значениям, непрерывные во времени.
Наиболее важными параметрами электронных сигналов являются следующие:
- Среднее значение сигнала ,
- Среднее значение сигнала в определенном диапазоне ,
- Энергия сигнала ,
- Мощность сигнала ,
- Средняя мощность сигнала в определенном диапазоне .
Синусоидальный сигнал (волна) – это непрерывная периодическая волна, описываемая двумя параметрами: амплитудой и частотой. Следующее уравнение описывает синусоидальный сигнал:
Если предположить, что ω = 2πf , то синусоидальный сигнал можно описать следующей формулой:
Иногда вместо амплитуды используют среднеквадратичное (СКЗ) значение напряжения размах напряжения В pp используется. Среднеквадратичное значение равно пиковому напряжению, умноженному на √2:
Полное значение равно удвоенному значению амплитуды В пиковое (пиковое напряжение):
Значения RMS используются при расчете энергии или мощности. При измерении электрического напряжения и силы тока мультиметры обычно показывают среднеквадратичное значение.
Рис. 3. Пример синусоиды Прямоугольный сигнал – описаниеПрямоугольный сигнал (волна) – аналогичен синусоиде, описывается двумя параметрами: амплитудой и частотой, с тем отличием, что среднеквадратичное напряжение значение для прямоугольной волны равно ее амплитуде. Частоту «f» часто заменяют периодом «T», равным:
Прямоугольный сигнал состоит из нарастающего фронта, высокого уровня, спадающего фронта и низкого уровня. Квадратные сигналы можно «обычно увидеть» только с положительными половинами, что означает, что низкий уровень сигнала близок к 0 В.
Прямоугольные сигналы используются как в цифровой электронике, так и в системах, граничащих с аналоговой электроникой, таких как компараторы, преобразователи АЦП (аналого-цифровой преобразователь) или ЦАП (цифро-аналоговый преобразователь).
В реальности форма волны отличается от идеальной формы склона, так как в реальности полностью перпендикулярным быть невозможно, но близко.
Рис. 4. Пример идеальной прямоугольной волныРис. 5. Пример реального прямоугольного сигнала Треугольный сигнал – ОписаниеТреугольный сигнал – треугольная волна, линейно нарастающая и падающая с определенной скоростью. Его можно получить (очень упрощенным способом) путем интегрирования прямоугольного сигнала.
Рис. 6. Пример треугольной волны Пилообразный сигнал – Описание Пилообразный сигнал (волна) – как следует из названия, напоминает форму зуба пилы. Этот сигнал имеет линейную форму – напряжение изменяется с фиксированной скоростью до определенного значения и периодически повторяется.
Шум – это неотъемлемая составляющая каждого сигнала, которая обычно нежелательна в электронных схемах.
Рис. 8. Пример шумового сигнала Импульсный сигнал – ОписаниеИмпульсные сигналы – большинство из них не являются периодическими сигналами. Они описываются двумя параметрами: амплитудой и шириной этих импульсов. Импульсные сигналы делятся на положительные и отрицательные. В случае цифровой электроники нам также потребуется анализировать периодические импульсы, которые далее описываются частотой и коэффициентом наполненности.
Рис. 9. Пример прямоугольного импульсаРис. 10. Примеры положительных импульсов Рис. 11. Примеры отрицательных импульсов Единичные ступени и пиковые сигналы – описание Единичные ступени и пиковые сигналы – можно констатировать, что эти сигналы практически не используются в электронных системах, но используются для их описания и анализа.
На следующем рисунке показан цифровой сигнал, состоящий из четырех 3-битных слов в двоичном коде.
Рис. 14. Пример цифрового сигнала – двоичный кодЭлектрические сигналы сердца
Электрическая система вашего сердца контролирует все события, которые происходят, когда ваше сердце качает кровь. Электрическая система также называется сердечной проводящей системой. Сердечный тест, называемый ЭКГ (электрокардиограмма), представляет собой графическое изображение электрической активности сердца.
Электрическая система вашего сердца состоит из трех основных частей:
- Синоатриальный (СА) узел, расположенный в правом предсердии вашего сердца.
- Атриовентрикулярный (АВ) узел, расположенный на межпредсердной перегородке рядом с трехстворчатым клапаном.
- Система Гиса-Пуркинье, расположенная вдоль стенок желудочков сердца.
Сердцебиение — это сложный цикл электропроводящих событий. Эти события происходят внутри и вокруг вашего сердца. Сердцебиение — это единый цикл, в котором камеры вашего сердца расслабляются и сокращаются, чтобы перекачивать кровь. Этот цикл включает в себя открытие и закрытие впускного и выпускного клапанов правого и левого желудочков сердца. Каждое сердцебиение состоит из двух основных частей: диастолы и систолы. Во время диастолы предсердия и желудочки вашего сердца расслабляются и начинают наполняться кровью.
В конце диастолы предсердия вашего сердца сокращаются (систола предсердий) и перекачивают кровь в желудочки. Затем предсердия начинают расслабляться. Затем желудочки сердца сокращаются (систола желудочков), выталкивая кровь из сердца.
Два разных типа клеток в вашем сердце позволяют электрическому сигналу контролировать ваше сердцебиение:
i. Проводящие клетки несут электрический сигнал вашего сердца.
ii. Мышечные клетки позволяют камерам вашего сердца сокращаться, и это действие запускается электрическим сигналом вашего сердца.
Электрический сигнал проходит через сеть проводящих клеточных «путей», которые стимулируют сокращение ваших верхних камер (предсердий) и нижних камер (желудочков). Сигнал может проходить по этим путям посредством сложной реакции, которая позволяет каждой клетке активировать соседнюю, стимулируя ее «пропускать» электрический сигнал упорядоченным образом. Поскольку клетка за клеткой быстро передает электрический заряд, все сердце сокращается одним скоординированным движением, создавая сердцебиение.
Проводящий цикл сердечного сокращения
(1) Синоатриальный (СА) узел и остальная часть проводящей системы находятся в состоянии покоя.
(2) Узел SA инициирует потенциал действия, который распространяется по предсердиям.
(3) После достижения атриовентрикулярного узла происходит задержка примерно 100 мс, которая позволяет предсердиям завершить перекачку крови до того, как импульс будет передан на атриовентрикулярный пучок.
(4) После задержки импульс проходит через атриовентрикулярный пучок и его ветви к волокнам Пуркинье, а также через модераторный пучок достигает правой сосочковой мышцы.
(5) Импульс распространяется на сократительные волокна желудочка.
(6) Начинается сокращение желудочков.
• Потенциал действия. Этот постепенный рост электрического потенциала называется пейсмекером или препотенциалом. Скорость нарастания пейсмекерного потенциала является основной детерминантой частоты сердечных сокращений и увеличивается при адреналиновой (эпинефрин) и симпатической стимуляции и снижается при вагусной (вызывающей падение кровяного давления) стимуляции и гипотермии. Активность кардиостимулятора обычно возникает только в СА и АВ узлах, но есть латентные водители ритма в других частях проводящей системы, которые вступают во владение, когда возбуждение из СА или АВ узлов подавлено. Мышечные волокна предсердий и желудочков не имеют кардиостимуляторной активности и спонтанно разряжаются только при повреждении или аномалиях.
Специализированная проводящая система сердца состоит из проводящей ткани, состоящей из модифицированных клеток сердечной мышцы, обладающих свойством автоматизма, то есть они могут генерировать собственные внутренние потенциалы действия (нервные импульсы), а также реагировать на стимуляцию соседних клеток . Проводящие пути внутри сердца ответственны за организованное распространение потенциалов действия в сердце и, как следствие, координированное сокращение как предсердий, так и желудочков. В ткани кардиостимулятора после реполяризации мембранный потенциал постепенно повышается до порогового уровня для открытия канала, после чего натрий поступает в клетку и инициирует следующий потенциал действия (нервный импульс).
Клетки СА узла в верхней части сердца известны как водители ритма сердца, потому что частота, с которой эти клетки посылают электрические сигналы, определяет частоту, с которой бьется все сердце (частоту сердечных сокращений). Нормальная частота сердечных сокращений в состоянии покоя колеблется от 60 до 100 ударов в минуту. Ваш сердечный ритм может повышаться или понижаться в соответствии с потребностями вашего организма. Ваш мозг и другие части вашего тела посылают сигналы, чтобы заставить ваше сердце биться быстрее или медленнее. Хотя способ взаимодействия всех химических сигналов, влияющих на частоту сердечных сокращений, сложен, конечным результатом является то, что эти сигналы сообщают узлу SA о необходимости запуска зарядов в более быстром или более медленном темпе, что приводит к ускорению или замедлению частоты сердечных сокращений.
Например, во время упражнений, когда организму требуется больше кислорода для функционирования, сигналы от вашего тела вызывают значительное увеличение частоты сердечных сокращений, чтобы доставить больше крови (и, следовательно, больше кислорода) в организм. Ваш сердечный ритм может увеличиться выше 100 ударов в минуту, чтобы удовлетворить повышенные потребности вашего тела во время физических нагрузок.
Точно так же в периоды отдыха или сна, когда организму требуется меньше кислорода, снижается частота сердечных сокращений. На самом деле у некоторых спортсменов нормальная частота сердечных сокращений может быть значительно ниже 60, потому что их сердце работает очень эффективно и ему не нужно биться так быстро. Таким образом, изменения частоты сердечных сокращений являются нормальной частью усилий вашего сердца, направленных на удовлетворение потребностей вашего тела.
Катехоламины. Во время стресса или потребности в увеличении сердечного выброса надпочечники выделяют в кровоток гормон норадреналин, в то время как симпатическая нервная система активируется для увеличения частоты сердечных сокращений. Этот гормон заставляет сердце биться быстрее, и в отличие от симпатической нервной системы, которая посылает мгновенный и кратковременный сигнал, норадреналин, выбрасываемый в кровоток, увеличивает частоту сердечных сокращений на несколько минут и более.
Графическая запись ЭКГ на бумаге
ЭКГ обычно записывается в масштабе времени 0,04 секунды/мм по горизонтальной оси и с вольтажной чувствительностью 0,1 мВ/мм по вертикальной оси.
Таким образом, на стандартной бумаге для записи ЭКГ 1 маленький квадрат соответствует 0,04 секунды, а один большой квадрат — 0,2 секунды.
На нормальной ЭКГ зубец P соответствует деполяризации предсердий, комплекс QRS — деполяризации желудочков, а зубец T — реполяризации желудочков. 915:15 Интервал P-R берется от начала зубца P до начала комплекса QRS. Интервал Q-T берется от начала комплекса QRS до окончания зубца Т. Это время, необходимое для деполяризации и реполяризации желудочков. Сегмент S-T представляет собой период между окончанием комплекса QRS и началом зубца Т. В норме все клетки в этой фазе деполяризованы.
Нормальные значения ЭКГ
Интервал P-R 0,12–0,2 секунды (3–5 маленьких квадратов стандартной бумаги для ЭКГ)
Продолжительность комплекса QRS меньше или равна 0,1 секунды (2,5 маленьких квадрата)
Интервал Q — T с поправкой на частоту сердечных сокращений (QTc) QTc = интервал QT/RR меньше или равен 0,44 секунды
Положения отведений
ЭКГ можно использовать двумя способами. Может быть выполнена ЭКГ в 12 отведениях, которая анализирует электрическую активность сердца с помощью нескольких электродов, расположенных на конечностях и поперек грудной клетки. Может быть обнаружен широкий спектр аномалий.
Однако во время анестезии ЭКГ отслеживается с использованием только 3 (иногда 5) электродов, которые обеспечивают более ограниченный анализ электрической активности сердца и не могут предоставить такой же объем информации, как ЭКГ в 12 отведениях.
Термин «отведение» применительно к ЭКГ не относится к электрическим кабелям, подсоединенным к электродам на пациенте. Вместо этого это относится к расположению двух электродов, используемых для обнаружения электрической активности сердца. Третий электрод действует как нейтральный.
Во время анестезии обычно используется одно из 3 возможных «отведений». Эти отведения называются биполярными отведениями, поскольку они измеряют разность потенциалов (электрическую разность) между двумя электродами. Электрическая активность, идущая к электроду, отображается на экране как положительное (вверх) отклонение, а электрическая активность, идущая в сторону, как отрицательное (вниз) отклонение. Отведения условно описываются следующим образом:
Отведение I — измеряет разность потенциалов между электродом правой руки и электродом левой руки. Третий электрод (левая нога) действует как нейтральный.
Отведение II — измеряет разность потенциалов между электродами на правой руке и левой ноге.
Отведение III — измеряет разность потенциалов между электродами на левой руке и левой ноге.
Большинство мониторов могут одновременно отображать только одно отведение, поэтому следует выбирать отведение, дающее максимальное количество информации. Наиболее часто используемым отведением является отведение II — биполярное отведение с электродами на правой руке и левой ноге. Это наиболее полезное отведение для выявления сердечных аритмий, поскольку оно расположено близко к оси сердца (общее направление электрического движения) и обеспечивает наилучший обзор зубцов P и R.
Источники:
деятельность
http://www.webmd.com/heart/tc/electrical-system-of-the-heart-topic-overview#1
https://www.nhlbi.nih.gov/health/health-topics/ топики/hhw/electrical
Составитель: John Sandham
электромеханический преобразователь | инструмент | Британика
электромеханический преобразователь
См. все материалы
- Связанные темы:
- преобразователь
Просмотреть весь связанный контент →
электромеханический преобразователь , устройство любого типа, которое либо преобразует электрический сигнал в звуковые волны (как в громкоговорителе), либо преобразует звуковую волну в электрический сигнал (как в микрофоне). Многие преобразователи, используемые в повседневной жизни, работают в обоих направлениях, например, спикерфон на некоторых домофонах.
Чтобы оценить пригодность различных типов микрофонов для конкретных целей, необходимо учитывать линейность их частотной характеристики, характеристики направленности, долговечность и стоимость.
Линейность и направленность
Частотная линейность — это способность микрофона выдавать электрический выходной сигнал, пропорциональный амплитуде входного звука во всем диапазоне частот. Для музыки это должно распространяться на гораздо более низкие и гораздо более высокие частоты, чем только для голоса. Для высококачественного музыкального воспроизведения лучше всего подходит более плоский или почти линейный микрофон.
Микрофоны также имеют характеристики направленности. Те, которые равномерно улавливают сигналы, поступающие со всех направлений, называются всенаправленными. Распространенным направленным микрофоном является кардиоидный микрофон, названный так потому, что, когда характеристика интенсивности как функция угла отображается на полярной диаграмме, кривая имеет форму сердца. Кардиоидный микрофон полезен для записи живых выступлений, где желательно устранить шум аудитории. Микрофон-пушка имеет очень сильную прямую направленную характеристику. Параболический рефлектор, похожий на телескоп-рефлектор, используется для улавливания и усиления относительно слабых звуков, исходящих с определенного направления. Это полезно для таких разнообразных приложений, как прослушивание криков птиц и прослушивание сигналов квотербека во время футбольного матча.
Типы датчиков
В большинстве микрофонов используется электромагнитный или электростатический метод преобразования звуковых волн в электрические сигналы. Динамический микрофон состоит из небольшого магнита, который колеблется внутри катушки, прикрепленной к диафрагме. Когда звуковая волна вызывает вибрацию диафрагмы микрофона, относительное движение магнита и катушки создает электрический сигнал за счет магнитной индукции. Можно использовать либо систему с подвижной катушкой, либо систему с подвижным магнитом, в зависимости от того, какой элемент соединен с подвижной диафрагмой; подвижная катушка используется чаще. Динамический микрофон прочный и имеет достаточно хорошую линейность, поэтому высококачественные модели полезны при записи. Поскольку микрофон с подвижной катушкой и громкоговоритель с подвижной катушкой очень похожи, переговорные устройства часто изготавливаются с одним и тем же элементом, выполняющим обе функции.
Электростатический или конденсаторный микрофон имеет диафрагму в виде одной пластины конденсатора с параллельными пластинами. Наиболее популярной формой микрофона этого типа является электретный конденсаторный микрофон, в котором пластины постоянно заряжаются электрическим током. Когда звуковая волна вызывает вибрацию заряженной пластины диафрагмы, напряжение на пластинах изменяется, создавая сигнал, который можно усилить и передать на записывающее устройство. Усилитель часто устанавливается в микрофон, поэтому этот тип микрофона требует использования батареи для питания усилителя. Поскольку диафрагма конденсаторного микрофона может быть очень легкой по сравнению с более массивным динамическим микрофоном, он способен реагировать быстрее и на более высоких частотах. Следовательно, конденсаторные микрофоны обычно имеют лучшую линейность и больший частотный диапазон, чем динамические микрофоны.
Оформите подписку Britannica Premium и получите доступ к эксклюзивному контенту. Подпишитесь сейчас
В кристаллическом микрофоне в качестве преобразователя используется пьезоэлектрический кристалл. Пьезоэлектрические кристаллы долговечны и дешевы, и они имеют относительно большую электрическую мощность; по этой причине они часто используются в телефонах и портативных звуковых системах. У них не очень хорошая линейность, поэтому они не подходят для качественной записи звука.
Ленточный микрофон уникален тем, что он реагирует на скорость звуковой волны, а не на изменение давления. Поскольку ленточные микрофоны очень чувствительны, их нельзя использовать там, где они могут подвергаться механическим ударам. Ленточные микрофоны являются двунаправленными и могут одинаково хорошо улавливать звуки, исходящие с обеих сторон микрофона.
Электромагнитные динамики
Большинство громкоговорителей относятся к электромагнитным, или динамическим, типам, в которых звуковая катушка перемещается в зазоре постоянного магнита, когда через катушку протекает переменный во времени ток. Магнит обычно имеет форму буквы «W» или кольца. Диафрагма или конус такого громкоговорителя движется вместе с катушкой, преобразуя электрический ток в катушке в волну давления. Зажженная свеча, помещенная перед диффузором громкоговорителя, который колеблется с частотой около 10 герц, может сделать звуковую волну «видимой», поскольку пламя колеблется в продольном направлении вместе с воздухом.
Как и в случае с микрофонами, громкоговорители оцениваются в основном по их частотной линейности. Чтобы добиться хорошей АЧХ на низких частотах, необходимо использовать довольно большой диффузор; однако из-за относительно большой массы катушки и диффузора громкоговорителя трудно добиться хорошего отклика на высоких частотах с тем же громкоговорителем. Отклик можно улучшить, используя довольно большие магниты, но они делают хороший громкоговоритель довольно тяжелым. Кроме того, критически важным является подвес катушки в зазоре магнита, поскольку он должен обеспечивать как быстродействие, так и быстрое затухание до положения равновесия при прекращении сигнала. Каждый громкоговоритель имеет частоту, на которой он резонирует наиболее легко. Для больших громкоговорителей эта резонансная частота полезна для усиления низких частот системы.
Громкоговорители монтируются в коробку, рупор или другой корпус, чтобы отделить волны от передней и задней части громкоговорителя и тем самым предотвратить их подавление друг друга. Наиболее распространенным типом корпуса является акустическая подвесная система, в которой громкоговоритель монтируется в герметичном корпусе. Для предотвращения резонансов в коробке типа, описываемого уравнением (36) в звуке изделия, внутренняя часть обычно покрыта каким-либо звукопоглощающим материалом. Из-за воздухонепроницаемого уплотнения конус должен сжимать и расширять воздух внутри коробки при движении, так что этот тип системы не очень эффективен в преобразовании электрической энергии в звук, особенно на низких частотах.
Корпус с настроенным портом или фазоинвертором обеспечивает большую эффективность и расширяет диапазон низких частот за счет тщательной регулировки формы и положения отверстия или трубки, соединяющей внутреннюю часть корпуса динамика с внешней стороной. Таким образом, объем ящика действует как своего рода резонатор Гельмгольца с резонансной частотой, которая определяется геометрией отверстия или трубы и намеренно выбрана таким образом, чтобы плавно расширять частотный диапазон акустической системы до значительно более низкого значения. . Кроме того, наличие порта значительно снижает колебания давления воздуха внутри корпуса, позволяя диффузору громкоговорителя двигаться более свободно. По этим причинам типичный корпус с фазоинвертором намного эффективнее, чем типичная система акустической подвески.
В корпусе рупора используется расширяющаяся трубка для получения наилучшей акустической связи между диффузором громкоговорителя и внешней средой, благодаря чему из диффузора громкоговорителя излучается наилучшая возможная когерентная волна. Такая система чрезвычайно эффективна и поэтому используется в системах громкой связи, театрах под открытым небом или других местах, где требуется большая акустическая мощность. Поскольку корпус басового рупора хорошего качества очень большой, в таких устройствах часто используются изогнутые или сложенные трубки. Klipschorn, названный в честь его изобретателя, американского инженера Пола В. Клипша, использует стены в углу комнаты как часть расширяющегося рупора.
Поскольку высокая эффективность и линейность одного динамика не могут быть распространены на весь диапазон слышимых частот, системы громкоговорителей обычно состоят из двух или более отдельных громкоговорителей. Динамик большего размера, или низкочастотный динамик, воспроизводит более низкие частоты, а динамик меньшего размера, или твитер, воспроизводит более высокие частоты. В такой двусторонней системе пассивная электронная схема, называемая перекрестной сетью, используется для направления высоких и низких частот на соответствующий громкоговоритель. Более крупная или более эффективная трехполосная система может добавить среднечастотный динамик, помогая создать более линейную характеристику между низкочастотным и высокочастотным динамиками.
Массивы громкоговорителей, которые обычно используются в больших залах, часто используют один низкочастотный и один среднечастотный динамик, но два или даже три высокочастотных твитера. Необходимость использования большего количества твитеров возникает из-за относительно меньшей дифракции высокочастотных (или низковолновых) звуковых волн. Поскольку они меньше распространяются и, следовательно, имеют более направленное звучание, может потребоваться несколько твитеров и направить их так, чтобы они покрывали всю аудиторию. Это не нужно для вуфера из-за большой дифракции длинных волн.
Электростатические динамики
В электростатических громкоговорителях используется большая тонкая металлическая пластина между двумя параллельными экранами. Усиленный звуковой сигнал попадает на экраны, поляризуя металлический лист, и результирующая электростатическая сила создает движение листа, создавая звуковую волну. Электростатические динамики хорошо работают на высоких частотах, но они не могут перемещать достаточно воздуха, чтобы хорошо работать на низких частотах, и часто требуют несколько большей мощности, чем электромагнитные динамики. Из-за этих ограничений и других технических проблем они нашли лишь ограниченное применение и не популярны в потребительских аудиосистемах.
Электрические сигналы, которые делают человеческий мозг уникальным
Время чтения:
Хотите БЕСПЛАТНУЮ версию этой новости в формате PDF?
Заполните форму ниже, и мы вышлем вам PDF-версию «Электрические сигналы, которые делают человеческий мозг уникальным»
Нейроны в человеческом мозгу получают электрические сигналы от тысяч других клеток, и длинные нервные отростки, называемые дендритами, играют решающую роль в объединении всей этой информации, чтобы клетки могли реагировать соответствующим образом.
Используя труднодоступные образцы ткани человеческого мозга, нейробиологи Массачусетского технологического института обнаружили, что электрические свойства дендритов человека отличаются от свойств дендритов других видов. Их исследования показывают, что электрические сигналы ослабевают больше, когда они проходят по дендритам человека, что приводит к более высокой степени электрической компартментализации, а это означает, что небольшие участки дендритов могут вести себя независимо от остальной части нейрона.
Эти различия могут способствовать повышению вычислительной мощности человеческого мозга, говорят исследователи.
«Люди умны не только потому, что у них больше нейронов и больше кора головного мозга. Снизу вверх нейроны ведут себя по-разному», — говорит Марк Харнетт, доцент Фреда и Кэрол Миддлтон по развитию карьеры в области мозга и когнитивных наук. «В нейронах человека электрическая компартментализация больше, и это позволяет этим единицам быть немного более независимыми, что потенциально приводит к увеличению вычислительных возможностей отдельных нейронов».
Харнетт, который также является членом Института исследований мозга Макговерна при Массачусетском технологическом институте, и Сидни Кэш, доцент кафедры неврологии Гарвардской медицинской школы и Массачусетской больницы общего профиля, являются ведущими авторами исследования, опубликованного 18 октября. выпуск Cell. Ведущим автором статьи является Лу Болье-Ларош, аспирант кафедры мозговых и когнитивных наук Массачусетского технологического института.
Нейронные вычисления
Дендриты можно рассматривать как аналог транзисторов в компьютере, выполняющих простые операции с использованием электрических сигналов. Дендриты получают информацию от многих других нейронов и передают эти сигналы в тело клетки. При достаточной стимуляции нейрон запускает потенциал действия — электрический импульс, который затем стимулирует другие нейроны. Большие сети этих нейронов взаимодействуют друг с другом, чтобы генерировать мысли и поведение.
Структура отдельного нейрона часто напоминает дерево с множеством ветвей, несущих информацию, поступающую далеко от тела клетки. Предыдущие исследования показали, что сила электрических сигналов, поступающих в тело клетки, частично зависит от того, как далеко они проходят по дендриту, чтобы попасть туда. По мере распространения сигналы становятся слабее, поэтому сигнал, приходящий далеко от тела клетки, оказывает меньшее влияние, чем сигнал, приходящий вблизи тела клетки.
Дендриты в коре головного мозга человека намного длиннее, чем у крыс и большинства других видов, потому что кора человека в процессе эволюции стала намного толще, чем у других видов. У человека кора головного мозга составляет около 75 процентов от общего объема мозга по сравнению с примерно 30 процентами в мозге крысы.
Хотя кора головного мозга человека в два-три раза толще, чем у крыс, она сохраняет ту же общую организацию, состоящую из шести различных слоев нейронов. Нейроны из слоя 5 имеют дендриты, достаточно длинные, чтобы дойти до слоя 1, а это означает, что человеческие дендриты должны были удлиняться по мере развития человеческого мозга, и электрические сигналы должны проходить гораздо дальше.
В новом исследовании команда Массачусетского технологического института хотела выяснить, как эти различия в длине могут повлиять на электрические свойства дендритов. Им удалось сравнить электрическую активность дендритов крысы и человека, используя небольшие кусочки мозговой ткани, взятые у пациентов с эпилепсией, перенесших хирургическое удаление части височной доли. Чтобы добраться до пораженной части мозга, хирурги также должны удалить небольшой кусок передней височной доли.
С помощью сотрудников MGH Кэша, Мэтью Фроша, Зива Уильямса и Эмада Эскандара лаборатория Харнетта смогла получить образцы передней височной доли, каждый размером с ноготь.
Имеющиеся данные свидетельствуют о том, что передняя височная доля не поражается эпилепсией, и ткань кажется нормальной при исследовании нейропатологическими методами, говорит Харнетт. Эта часть мозга, по-видимому, участвует во множестве функций, включая язык и визуальную обработку, но не имеет решающего значения ни для одной функции; пациенты могут нормально функционировать после его удаления.
После того, как ткань была удалена, исследователи поместили ее в раствор, очень похожий на спинномозговую жидкость, через который циркулировал кислород. Это позволило им сохранить ткани живыми до 48 часов. В то время они использовали метод, известный как электрофизиология пэтч-кламп, для измерения того, как электрические сигналы проходят по дендритам пирамидальных нейронов, которые являются наиболее распространенным типом возбуждающих нейронов в коре.
Эти эксперименты были выполнены прежде всего Beaulieu-Laroche. Лаборатория Харнетта (и другие) ранее проводили подобные эксперименты на дендритах грызунов, но его команда первой проанализировала электрические свойства дендритов человека.
Уникальные особенности
Исследователи обнаружили, что, поскольку дендриты человека покрывают большие расстояния, сигнал, проходящий по дендриту человека от слоя 1 к телу клетки в слое 5, когда приходит, намного слабее, чем сигнал, проходящий по дендриту крысы от слоя 1 на слой 5.
Они также показали, что дендриты человека и крысы имеют одинаковое количество ионных каналов, которые регулируют ток, но эти каналы имеют более низкую плотность в дендритах человека в результате удлинения дендритов. Они также разработали подробную биофизическую модель, которая показывает, что это изменение плотности может объяснить некоторые различия в электрической активности, наблюдаемые между дендритами человека и крысы, говорит Харнетт.
Нельсон Спрустон, старший директор научных программ в Исследовательском кампусе Джанелии Медицинского института Говарда Хьюза, назвал проведенный исследователями анализ человеческих дендритов «выдающимся достижением».
«На сегодняшний день это самые подробные измерения физиологических свойств нейронов человека», — говорит Спрустон, не участвовавший в исследовании. «Такого рода эксперименты очень сложны с технической точки зрения, даже на мышах и крысах, поэтому с технической точки зрения удивительно, что они сделали это на людях».
Остается вопрос, как эти различия влияют на возможности человеческого мозга? Гипотеза Харнетта заключается в том, что из-за этих различий, которые позволяют большему количеству областей дендрита влиять на силу входящего сигнала, отдельные нейроны могут выполнять более сложные вычисления с информацией.
«Если у вас есть кортикальная колонка с фрагментом коры головного мозга человека или грызуна, вы сможете выполнять больше вычислений быстрее с человеческой архитектурой по сравнению с архитектурой грызунов», — говорит он.