
Действие постоянного тока на возбудимые ткани
Впервые
закономерности действия постоянного
тока на нерв нервно-мышечного препарата
исследовал в 19 веке Пфлюгер. Он установил,
что при замыкании цепи постоянного
тока, под отрицательным электродом,
т.е. катодом возбудимость повышается,
а под положительным — анодом снижается.
Это называется законом действия
постоянного тока. Изменение возбудимости
ткани (например нерва) под действием
постоянного тока в области анода или
катода называется физиологическим
электротоном. В настоящее время
установлено, что под действием
отрицательного электрода — катода
потенциал мембраны клеток снижается.
Это явление называется физическим
катэлектротоном. Под положительным —
анодом, он возрастает. Возникает
физический анэлектртон. Так как, под
катодом мембранный потенциал приближается
к критическому уровню деполяризации,
возбудимость клеток и тканей повышается.
Под анодом мембранный потенциал
возрастает и удаляется от критического
уровня деполяризации, поэтому возбудимость
клетки, ткани падает.
Постоянный ток широко используется в клинике для лечения и диагностики. Например, с помощью него производится электростимуляция нервов и мышц, физиопроцедуры: ионофорез и гальванизация.
Строение и функции цитоплазматической мембраны клеток.
Цитоплазматическая
клеточная мембрана состоит из трех
слоев: наружного белкового, среднего
бимолекулярного слоя липидов и внутреннего
белкового. Толщина мембраны 7,5-10 нМ.
Бимолекулярный слой липидов является
матриксом мембраны. Липидные молекулы
его обоих слоев взаимодействуют с
белковыми молекулами, погруженными в
них. От 60 до 75% липидов мембраны составляют
фосфолипиды, 15-30% холестерин. Белки
представлены в основном гликопротеинами.
Различают интегральные белки, пронизывающие
всю мембрану и периферические, находящиеся
на наружной или внутренней поверхности.
Функции мембраны:
Обеспечивает целостность клетки, как структурной единицы ткани.
Осуществляет обмен ионов между цитоплазмой и внеклеточной жидкостью.
3. Обеспечивает активный транспорт ионов и других веществ в клетку и из нее
4. Производит восприятие и переработку информации поступающей к клетке в виде химических и электрических сигналов.
Механизмы возбудимости клеток. Ионные каналы мембраны. Механизмы возникновения мембранного потенциала (мп) и потенциалов действия (пд)
В основном,
передаваемая в организме информация
имеет вид электрических сигналов
(например нервные импульсы).

Законы действия постоянного тока на возбудимые ткани
Полярный закон действия тока. При раздражении нерва или мышцы постоянным током возбуждение возникает в момент замыкания постоянного тока только под катодом, а в момент размыкания — только под анодом, причем порог замыкательного удара меньше, чем размыкательного. Прямые измерения показали, что прохождение через нервное или мышечное волокно электрического тока вызывает прежде всего изменение мембранного потенциала под электродами. В области приложения к поверхности ткани анода (+) положительный потенциал на наружной поверхности мембраны возрастает, т.е. в этом участке происходит гиперполяризация мембраны, что не способствует возбуждению, а, наоборот, ему препятствует. В том же участке, где к мембране приложен катод (-), положительный потенциал наружной поверхности снижается, происходит деполяризация, и если она достигает критической величины — в этом месте возникает ПД.
В области приложения к поверхности ткани анода (+) положительный потенциал на наружной поверхности мембраны возрастает, т.е. в этом участке происходит гиперполяризация мембраны, что не способствует возбуждению, а, наоборот, ему препятствует. В том же участке, где к мембране приложен катод (-), положительный потенциал наружной поверхности снижается, происходит деполяризация, и если она достигает критической величины — в этом месте возникает ПД.
Изменение МП возникают не только непосредственно в точках приложения к нервному волокну катода и анода, но и на некотором расстоянии от них, но величина этих сдвигов убывает по мере удаления от электродов. Изменения МП под электродами носят название электротонических ( соответственно кат-электротон и ан-электротон), а за электродами — периэлектротонических (кат- и ан-периэлектротон).
Увеличение МП под анодом (пассивная гиперполяризация) не сопровождается изменением ионной проницаемости мембраны даже при большой силе приложенного тока. Поэтому при замыкании постоянного тока возбуждение под анодом не возникает. В отличие от этого, уменьшение МП под катодом (пассивная деполяризация) влечет за собой кратковременное повышение проницаемости для Na, что приводит к возбуждению.
Поэтому при замыкании постоянного тока возбуждение под анодом не возникает. В отличие от этого, уменьшение МП под катодом (пассивная деполяризация) влечет за собой кратковременное повышение проницаемости для Na, что приводит к возбуждению.
Повышение проницаемости мембраны для Na при пороговом раздражении не сразу достигает максимальной величины. В первый момент деполяризация мембраны под катодом приводит к небольшому увеличению натриевой проницаемости и открытию небольшого числа каналов. Когда же под влиянием этого в протоплазму начинают поступать заряженные положительно ионы Na+, то деполяризация мембраны усиливается. Это ведет к открытию других Na-каналов, и, следовательно, к дальнейшей деполяризации, которая, в свою очередь, обуславливает еще большее повышение натриевой проницаемости. Этот круговой процесс, основанный на т.н. положительной обратной связи, получил название регенеративной деполяризации. Возникает она только при снижении Ео до критического уровня (Ек ). Причина повышения натриевой проницаемости при деполяризации связана, вероятно, с удалением Са++ из натриевых ворот при возникновении электро отрицательности (или снижении электро положительности) на наружной стороне мембраны.
Причина повышения натриевой проницаемости при деполяризации связана, вероятно, с удалением Са++ из натриевых ворот при возникновении электро отрицательности (или снижении электро положительности) на наружной стороне мембраны.
Повышенная натриевая проницаемость через десятые доли миллисекунды за счет механизмов натриевой инактивации прекращается.
Скорость, с которой происходит деполяризация мембраны, зависит от силы раздражающего тока. При слабой силе деполяризация развивается медленно, и поэтому для возникновения ПД такой стимул должен иметь большую длительность.
Локальный ответ, который возникает при под пороговых стимулах, так же, как и ПД, обусловлен повышением натриевой проницаемости мембраны. Однако при под пороговом стимуле это повышение недостаточно велико для того, чтобы вызвать процесс регенеративной деполяризации мембраны. Поэтому начавшаяся деполяризация приостанавливается инактивацией и повышением калиевой проницаемости.
Подводя итог изложенному выше, можно следующим образом изобразить цепь событий, развивающихся в нервном или мышечном волокне под катодом раздражающего тока: пассивная деполяризация мембраны —- повышение натриевой проницаемости — усиление потока Na внутрь волокна — активная деполяризация мембраны — локальный ответ — превышение Ек — регенеративная деполяризация — потенциал действия (ПД).
Каков же механизм возникновения возбуждения под анодом при размыкании? В момент включения тока под анодом мембранный потенциал возрастает — происходит гиперполяризация. При этом разница между Ео и Ек растет, и для того, чтобы сдвинуть МП до критического уровня, нужна большая сила. При выключении тока (размыкание) исходный уровень Ео восстанавливается. Казалось бы, в это время нет условий для возникновения возбуждения. Но это справедливо только для того случая, если действие тока продолжалось очень короткое время (менее 100 мсек.). При длительном действии тока начинает меняться сам критический уровень деполяризации — он растет. И, наконец, возникает момент, когда новый Ек становится равным старому уровню Ео. Теперь при выключении тока возникают условия для возбуждения, ибо мембранный потенциал становится равным новому критическому уровню деполяризации. Величина ПД при размыкании всегда больше, чем при замыкании.
Зависимость пороговой силы стимула от его длительности. Как уже указывалось, пороговая сила любого стимула в определенных пределах находится в обратной зависимости от его длительности. В особенно четкой форме эта зависимость проявляется при использовании в качестве раздражителя прямоугольных толчков постоянного тока. Полученная в таких опытах кривая получила название «кривой силы-времени». Она была изучена Гоорвегом, Вейсом и Лапиком в начале века. Из рассмотрения этой кривой прежде всего следует, что ток ниже некоторой минимальной величины или напряжения не вызывает возбуждения, как бы длительно он не действовал. Минимальная сила тока, способная вызвать возбуждение, названа Лапиком реобазой. Наименьшее время, в течение которого должен действовать раздражающий стимул, называют полезным временем. Усиление тока приводит к укорочению минимального времени раздражения, но не беспредельно. При очень коротких стимулах кривая силы-времени становится параллельной оси координат. Это означает, что при таких кратковременных раздражениях возбуждения не возникает, как бы ни была велика сила раздражения.
Как уже указывалось, пороговая сила любого стимула в определенных пределах находится в обратной зависимости от его длительности. В особенно четкой форме эта зависимость проявляется при использовании в качестве раздражителя прямоугольных толчков постоянного тока. Полученная в таких опытах кривая получила название «кривой силы-времени». Она была изучена Гоорвегом, Вейсом и Лапиком в начале века. Из рассмотрения этой кривой прежде всего следует, что ток ниже некоторой минимальной величины или напряжения не вызывает возбуждения, как бы длительно он не действовал. Минимальная сила тока, способная вызвать возбуждение, названа Лапиком реобазой. Наименьшее время, в течение которого должен действовать раздражающий стимул, называют полезным временем. Усиление тока приводит к укорочению минимального времени раздражения, но не беспредельно. При очень коротких стимулах кривая силы-времени становится параллельной оси координат. Это означает, что при таких кратковременных раздражениях возбуждения не возникает, как бы ни была велика сила раздражения.
Определение полезного времени практически затруднено, так как точка полезного времени находится на участке кривой, переходящей в параллельную. Поэтому Лапик предложил использовать полезное время двух реобаз — хронаксию. Ее точка находится на самом крутом участке кривой Гоорвега-Вейса. Хронаксиметрия получила широкое распространение как в эксперименте, так и в клинике для диагностики повреждений волокон двигательных нервов.
Зависимость порога от крутизны нарастания силы раздражителя. Величина порога раздражения нерва или мышцы зависит не только от длительности стимула, но и от крутизны нарастания его силы. Порог раздражения имеет наименьшую величину при толчках тока прямоугольной формы, характеризующихся максимально быстрым нарастанием тока. Если же вместо таких стимулов применять линейно или экспоненциально нарастающие стимулы, пороги оказываются увеличенными и тем больше, чем медленнее нарастает ток. При уменьшении крутизны нарастания тока ниже некоторой минимальной величины (т. н. критический наклон) ПД вообще не возникает, до какой бы конечной силы не увеличивался ток.
н. критический наклон) ПД вообще не возникает, до какой бы конечной силы не увеличивался ток.
Такое явление приспособления возбудимой ткани к медленно нарастающему раздражителю получило название аккомодация. Чем выше скорость аккомодации, тем более круто должен нарастать стимул, чтобы не утратить своего раздражающего действия. Аккомодация к медленно нарастающему току обусловлена тем, что за время действия этого тока в мембране успевают развиться процессы, препятствующие возникновению ПД.
Выше уже указывалось, что деполяризация мембраны приводит к началу двух процессов: одного — быстрого, ведущего к повышению натриевой проницаемости и возникновению ПД, а другого — медленного, приводящего к инактивации натриевой проницаемости и к окончанию возбуждения. При крутом нарастании стимула Na-активация успевает достичь значительной величины прежде, чем развивается Na-инактивация. В случае медленного нарастания силы тока на первый план выступают процессы инактивации, приводящие к повышению порога и снижению амплитуды ПД.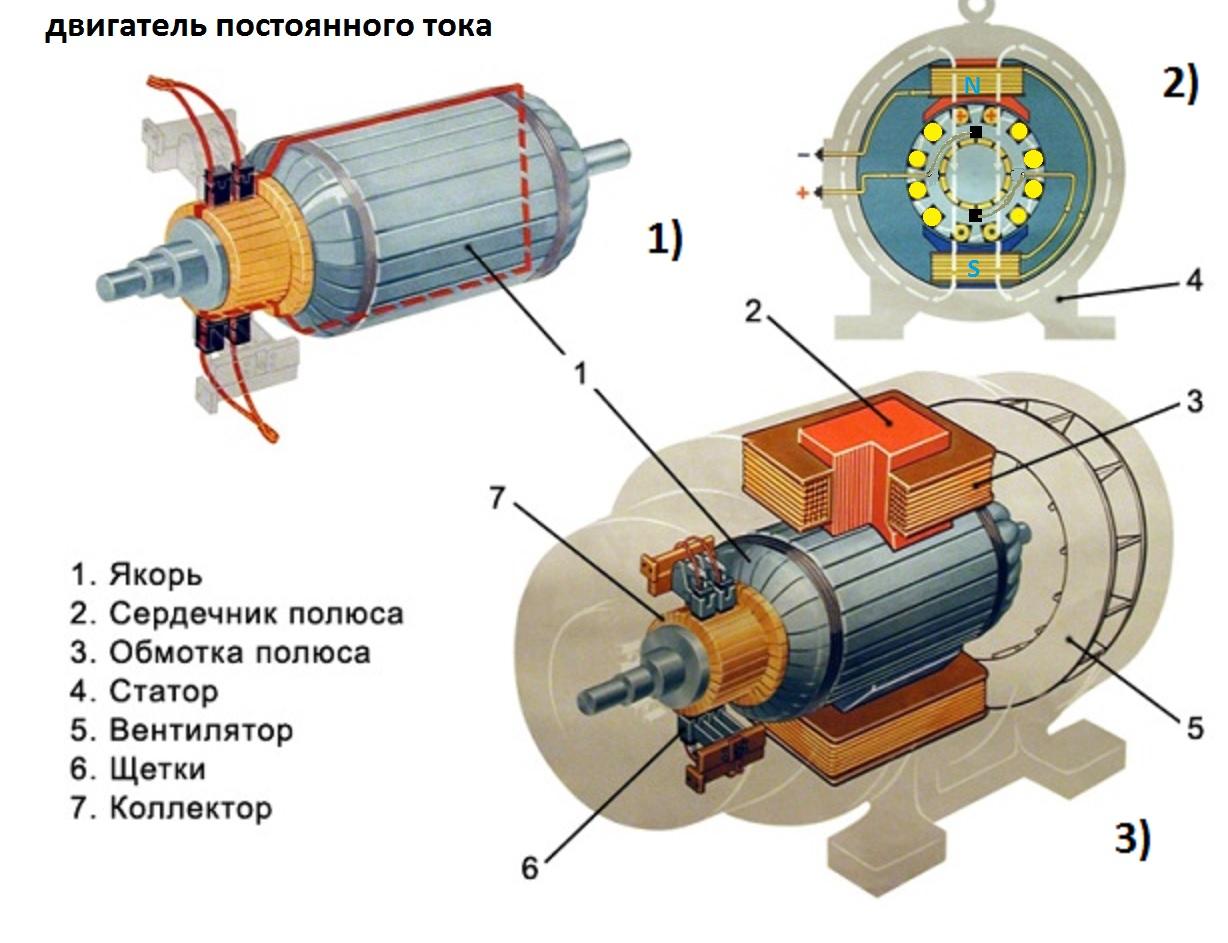 Все агенты, усиливающие или ускоряющие инактивацию, увеличивают скорость аккомодации.
Все агенты, усиливающие или ускоряющие инактивацию, увеличивают скорость аккомодации.
Аккомодация развивается не только при раздражении возбудимых тканей электрическим током, но и в случае применения механических, термических и прочих стимулов. Так, быстрый удар палочкой по нерву вызывает его возбуждение, при медленном же надавливании на нерв той же палочкой возбуждения не возникает. Изолированное нервное волокно можно возбудить быстрым охлаждением, а медленным — нельзя. Лягушка выпрыгнет, если ее бросить в воду с температурой 40 градусов, но если ту же лягушку поместить в холодную воду, и медленно нагревать ее, то животное сварится, но не будет реагировать прыжком на подъем температуры.
В лаборатории показателем скорости аккомодации является та наименьшая крутизна нарастания тока, при которой стимул еще сохраняет способность вызывать ПД. Эту минимальную крутизну называют критическим наклоном. Его выражают или в абсолютных единицах (мА/сек), или в относительных (как отношение пороговой силы того постепенно нарастающего тока, который еще способен вызывать возбуждение, к реобазе прямоугольного толчка тока).
Закон «все или ничего». При изучении зависимости эффектов раздражения от силы приложенного стимула был установлен т.н. закон «все или ничего». Согласно этому закону, под пороговые раздражения не вызывают возбуждения («ничего»), при пороговых же стимулах возбуждение сразу приобретает максимальную величину («все»), и уже не возрастает при дальнейшем усилении раздражителя.
Эта закономерность первоначально была открыта Боудичем при исследовании сердца, а в дальнейшем подтверждена и на других возбудимых тканях. Долгое время закон «все или ничего» неправильно интерпретировался как общий принцип реагирования возбудимых тканей. Предполагали, что «ничего» означает полное отсутствие ответа на под пороговый стимул, а «все» рассматривалось как проявление полного исчерпания возбудимым субстратом его потенциальных возможностей. Дальнейшие исследования, в особенности микроэлектродные, показали, что эта точка зрения не соответствует действительности. Выяснилось, что при под пороговых силах возникает местное не распространяющееся возбуждение (локальный ответ). Вместе с тем оказалось, что «все» также не характеризует того максимума, которого может достигнуть ПД. В живой клетке существуют процессы, активно приостанавливающие деполяризацию мембраны. Если каким-либо воздействием на нервное волокно, например, наркотиками, ядами, ослабить входящий Na-ток, обеспечивающий генерацию ПД, то он перестает подчиняться правилу «все или ничего» — его амплитуда начинает градуально зависеть от силы стимула. Поэтому «все или ничего» рассматривается сейчас не как всеобщий закон реагирования возбудимого субстрата на раздражитель, а лишь как правило, характеризующее особенности возникновения ПД в данных конкретных условиях.
Вместе с тем оказалось, что «все» также не характеризует того максимума, которого может достигнуть ПД. В живой клетке существуют процессы, активно приостанавливающие деполяризацию мембраны. Если каким-либо воздействием на нервное волокно, например, наркотиками, ядами, ослабить входящий Na-ток, обеспечивающий генерацию ПД, то он перестает подчиняться правилу «все или ничего» — его амплитуда начинает градуально зависеть от силы стимула. Поэтому «все или ничего» рассматривается сейчас не как всеобщий закон реагирования возбудимого субстрата на раздражитель, а лишь как правило, характеризующее особенности возникновения ПД в данных конкретных условиях.
Понятие о возбудимости. Изменения возбудимости при возбуждении.
Параметры возбудимости.
Возбудимостью называется способность нервной или мышечной клетки отвечать на раздражение генерацией ПД. Основным мерилом возбудимости обычно служит реобаза. Чем она ниже, тем выше возбудимость, и наоборот. Связано это с тем, что, как мы уже говорили ранее, главным условием возникновения возбуждения является достижение МП критического уровня деполяризации (Ео <= Ек). Поэтому мерилом возбудимости является разница между этими величинами (Ео — Ек). Чем меньше эта разница, тем меньшую силу надо приложить к клетке, чтобы сдвинуть мембранный потенциал до критического уровня, и, следовательно, тем больше возбудимость клетки.
Чем она ниже, тем выше возбудимость, и наоборот. Связано это с тем, что, как мы уже говорили ранее, главным условием возникновения возбуждения является достижение МП критического уровня деполяризации (Ео <= Ек). Поэтому мерилом возбудимости является разница между этими величинами (Ео — Ек). Чем меньше эта разница, тем меньшую силу надо приложить к клетке, чтобы сдвинуть мембранный потенциал до критического уровня, и, следовательно, тем больше возбудимость клетки.
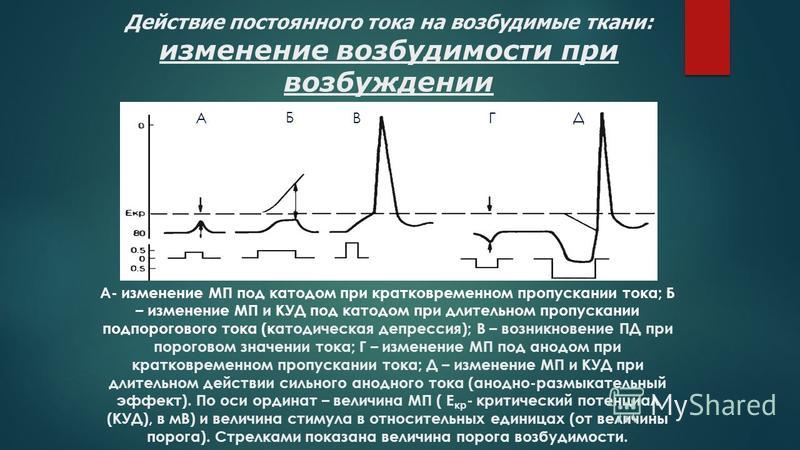
Еще Пфлюгер показал, что возбудимость — величина непостоянная. Катод повышает возбудимость, анод ее понижает. Напомним, что эти изменения возбудимости под электродами носят название электротонических. Русский ученый Вериго показал, что при длительном действии постоянного тока на ткань, или при действии сильных раздражителей эти электротонические изменения возбудимости извращаются — под катодом начальное повышение возбудимости сменяется ее понижением (развивается т.н. катодическая депрессия), а под анодом сниженная возбудимость постепенно возрастает. Причина этих изменений возбудимости на полюсах постоянного тока связана с тем, что величина Ек меняется при длительном действии раздражителя. Под катодом (и при возбуждении) Ек постепенно удаляется от МП, снижается, так что наступает момент, когда разница Е0-Ек становиться больше исходной. Это приводит к падению возбудимости ткани. Напротив, под анодом Ек имеет тенденцию к возрастанию, постепенно приближаясь к Ео. Возбудимость при этом растет, так как уменьшается исходная разница между Ео и Ек.
Причина этих изменений возбудимости на полюсах постоянного тока связана с тем, что величина Ек меняется при длительном действии раздражителя. Под катодом (и при возбуждении) Ек постепенно удаляется от МП, снижается, так что наступает момент, когда разница Е0-Ек становиться больше исходной. Это приводит к падению возбудимости ткани. Напротив, под анодом Ек имеет тенденцию к возрастанию, постепенно приближаясь к Ео. Возбудимость при этом растет, так как уменьшается исходная разница между Ео и Ек.
Причиной изменения критического уровня деполяризации под катодом является инактивация натриевой проницаемости, обусловленная длительной деполяризацией мембраны. Вместе с этим значительно повышается проницаемость для К. Все это приводит к тому, что мембрана клетки утрачивает способность отвечать на действие раздражающих стимулов. Такие же изменения мембраны лежат и в основе рассмотренного уже явления аккомодации. Под анодом же при действии тока явления инактивации снижаются .
Изменения возбудимости при возбуждении. Возникновение в нервном или мышечном волокне ПД сопровождается многофазными изменениями возбудимости. Для их изучения нерв или мышцу подвергают действию двух коротких электрических стимулов, следующих друг за другом с определенным интервалом. Первый называется раздражающим, второй — тестирующим. Регистрация возникающих в ответ на эти раздражения ПД позволила установить важные факты.
Возникновение в нервном или мышечном волокне ПД сопровождается многофазными изменениями возбудимости. Для их изучения нерв или мышцу подвергают действию двух коротких электрических стимулов, следующих друг за другом с определенным интервалом. Первый называется раздражающим, второй — тестирующим. Регистрация возникающих в ответ на эти раздражения ПД позволила установить важные факты.
Во время локального ответа возбудимость повышена, так как мембрана деполяризована и разность между Е0 и Ек падает. Периоду же возникновения и развития пика потенциала действия соответствует полное исчезновение возбудимости, получившее название абсолютной рефрактерности (невпечатлительности). В это время тестирующий стимул не способен вызвать новый ПД, как бы сильно ни было это раздражение. Длительность абсолютной рефрактерности примерно совпадает с длительностью восходящей ветви ПД. В быстро проводящих нервных волокнах она составляет 0,4-0,7 мсек. В волокнах мышцы сердца — 250-300 мсек. Вслед за абсолютной рефрактерностью начинается фаза относительной рефрактерности , которая длится 4-8 мсек. Она совпадает с фазой реполяризации ПД. В это время возбудимость постепенно возвращается к первоначальному уровню. В этот период нервное волокно способно ответить на сильное раздражение, но амплитуда ПД будет резко снижена.
Вслед за абсолютной рефрактерностью начинается фаза относительной рефрактерности , которая длится 4-8 мсек. Она совпадает с фазой реполяризации ПД. В это время возбудимость постепенно возвращается к первоначальному уровню. В этот период нервное волокно способно ответить на сильное раздражение, но амплитуда ПД будет резко снижена.
Согласно ионной теории Ходжкина-Хаксли, абсолютная рефрактерность обусловлена вначале наличием максимальной натриевой проницаемости, когда новый стимул не может что-то изменить или добавить, а затем развитием натриевой инактивации, закрывающей Na-каналы. Вслед за этим происходит снижение натриевой инактивации, в результате чего постепенно восстанавливается способность волокна генерировать ПД. Это — состояние относительной рефрактерности.
Относительная рефрактерная фаза сменяется фазой повышенной (супернормальной) возбудимости, совпадающей по времени с периодом следовой деполяризации. В это время разность между Ео и Ек ниже исходной. В двигательных нервных волокнах теплокровных животных длительность супернормальной фазы составляет 12-30 мсек.
В это время разность между Ео и Ек ниже исходной. В двигательных нервных волокнах теплокровных животных длительность супернормальной фазы составляет 12-30 мсек.
Период повышенной возбудимости сменяется субнормальной фазой , которая совпадает со следовой гиперполяризацией. В это время разница между мембранным потенциалом (Ео) и критическим уровнем деполяризации (Ек) увеличивается. Длительность этой фазы составляет несколько десятков или сотен мсек.
Лабильность. Мы рассмотрели основные механизмы возникновения и распространения в нервных и мышечных волокнах одиночной волны возбуждения. Однако в естественных условиях существования организма по нервным волокнам проходят не одиночные, а ритмические залпы потенциалов действия. В чувствительных нервных окончаниях, расположенных в любых тканях, возникают и распространяются по отходящим от них афферентным нервным волокнам ритмические разряды импульсов даже при очень кратковременном раздражении. Равным образом из ЦНС по эфферентным нервам идет поток импульсов на периферию к исполнительным органам. Если исполнительным органом являются скелетные мышцы, то в них возникают вспышки возбуждений в ритме поступающих по нерву импульсов.
Равным образом из ЦНС по эфферентным нервам идет поток импульсов на периферию к исполнительным органам. Если исполнительным органом являются скелетные мышцы, то в них возникают вспышки возбуждений в ритме поступающих по нерву импульсов.
Частота разрядов импульсов в возбудимых тканях может варьировать в широких пределах в зависимости от силы приложенного раздражения, свойств и состояния ткани и от скорости протекания отдельных актов возбуждения в ритмическом ряду. Для характеристики этой скорости Н.Е.Введенским и было сформулировано понятие лабильность. Под лабильностью, или функциональной подвижностью он понимал большую или меньшую скорость протекания тех элементарных реакций, которыми сопровождается возбуждение. Мерой лабильности является наибольшее число потенциалов действия, которое возбудимый субстрат способен воспроизвести в единицу времени в соответствии с частотой подаваемого раздражения.
Первоначально предполагали, что минимальный интервал между импульсами в ритмическом ряду должен соответствовать длительности абсолютного рефрактерного периода. Точные исследования, однако, показали, что при частоте следования стимулов с таким интервалом возникают только два импульса, а третий выпадает вследствие развивающейся депрессии. Поэтому интервал между импульсами должен быть несколько больше абсолютного рефрактерного периода. В двигательных нервных клетках теплокровных животных рефрактерный период составляет около 0,4 мсек, и потенциальный максимальный ритм должен бы быть равным 2500/сек, но на самом деле он равен около 1000/сек. Следует подчеркнуть, что эта частота значительно превышает частоту импульсов, проходящих по этим волокнам в физиологических условиях. Последняя составляет около 100/сек.
Точные исследования, однако, показали, что при частоте следования стимулов с таким интервалом возникают только два импульса, а третий выпадает вследствие развивающейся депрессии. Поэтому интервал между импульсами должен быть несколько больше абсолютного рефрактерного периода. В двигательных нервных клетках теплокровных животных рефрактерный период составляет около 0,4 мсек, и потенциальный максимальный ритм должен бы быть равным 2500/сек, но на самом деле он равен около 1000/сек. Следует подчеркнуть, что эта частота значительно превышает частоту импульсов, проходящих по этим волокнам в физиологических условиях. Последняя составляет около 100/сек.
Дело в том, что обычно в естественных условиях ткань работает с так называемым оптимальным ритмом. Для пропускания импульсов с таким ритмом не требуется большой силы раздражения. Исследования показали, что частота раздражения и реобаза тока, способного вызывать нервные импульсы с такой частотой, находятся в своеобразной зависимости: реобаза вначале падает по мере роста частоты импульсов, затем снова нарастает. Оптимум находится у нервов в пределах от 75 до 150 имп/сек, для мышц — 20-50 имп/сек. Такой ритм, в отличие от других, очень стойко и длительно может воспроизводиться возбудимыми образованиями.
Оптимум находится у нервов в пределах от 75 до 150 имп/сек, для мышц — 20-50 имп/сек. Такой ритм, в отличие от других, очень стойко и длительно может воспроизводиться возбудимыми образованиями.
Таким образом, мы можем теперь назвать все основные параметры возбудимости тканей, характеризующие ее свойства: РЕОБАЗА, ПОЛЕЗНОЕ ВРЕМЯ (ХРОНАКСИЯ), КРИТИЧЕСКИЙ НАКЛОН, ЛАБИЛЬНОСТЬ. Все они, кроме последнего, находятся с возбудимостью в обратно пропорциональных отношениях.
Понятие о «парабиозе«. Лабильность — величина непостоянная. Она может меняться в зависимости от состояния нерва или мышцы, в зависимости от силы и длительности падающих на них раздражений, от степени утомления и т.п. Впервые изменение лабильности нерва при действии на него сначала химическими, а затем и электрическими раздражителями, изучил Н.Е.Введенский. Он обнаружил закономерное снижение лабильности альтерированного химическим агентом (аммиаком) участка нерва, назвал это явление «парабиозом» и изучил его закономерности. Парабиоз — это обратимое состояние, которое, однако, при углублении действия вызывающего его агента может перейти в необратимое.
Парабиоз — это обратимое состояние, которое, однако, при углублении действия вызывающего его агента может перейти в необратимое.
Введенский рассматривал парабиоз как особое состояние стойкого не колеблющегося возбуждения, как бы застывшего в одном участке нервного волокна. Действительно, парабиотический участок заряжен отрицательно. Такое явление Введенский считал прообразом перехода возбуждения в торможение в нервных центрах. По его мнению, парабиоз является результатом перевозбуждения нервной клетки слишком сильным или слишком частым раздражением.
Развитие парабиоза протекает в три стадии: уравнительную, парадоксальную и тормозную. Вначале за счет снижения аккомодации отдельные импульсы тока малой частоту при условии их достаточной силы дают уже не 1 импульс, а 2,3 или даже 4. Одновременно порог возбудимости растет, а максимальный ритм возбуждения прогрессивно снижается. В результате на импульсы как малой, так и большой частоты нерв начинает отвечать одной и той же частотой разрядов, наиболее близкой к оптимальному для этого нерва ритму. Это и есть уравнительная фаза парабиоза. На следующем этапе развития процесса в области пороговых интенсивностей раздражения еще сохраняется воспроизведение ритма, близкого к оптимальному, а на частые импульсы ткань или вообще не отвечает, или отвечает очень редкими волнами возбуждения. Это — парадоксальная фаза.
Это и есть уравнительная фаза парабиоза. На следующем этапе развития процесса в области пороговых интенсивностей раздражения еще сохраняется воспроизведение ритма, близкого к оптимальному, а на частые импульсы ткань или вообще не отвечает, или отвечает очень редкими волнами возбуждения. Это — парадоксальная фаза.
Затем способность волокна к ритмической волновой деятельности падает, падает и амплитуда ПД, увеличивается его длительность., Любое внешнее воздействие подкрепляет состояние торможения нервного волокна и одновременно затормаживается само. Это — последняя, тормозная фаза парабиоза.
В настоящее время описанный феномен объясняется с позиций мембранной теории нарушением механизма повышения натриевой проницаемости и появлением затяжной натриевой инактивации. В результате этого Na-каналы остаются закрытыми, он накапливается в клетке и наружная поверхность мембраны длительное время сохраняет отрицательный заряд. Это препятствует новому раздражению за счет удлинения рефрактерного периода. При набегании на участок парабиоза часто следующих друг за другом ПД, инактивация натриевой проницаемости, вызванная альтерирующим агентом, суммируется с инактивацией, сопровождающей нервный импульс. В результате возбудимость снижается настолько, что проведение следующего импульса полностью блокируется.
Это препятствует новому раздражению за счет удлинения рефрактерного периода. При набегании на участок парабиоза часто следующих друг за другом ПД, инактивация натриевой проницаемости, вызванная альтерирующим агентом, суммируется с инактивацией, сопровождающей нервный импульс. В результате возбудимость снижается настолько, что проведение следующего импульса полностью блокируется.
Обмен веществ и энергии при возбуждении. При возникновении и проведении возбуждения в нервных клетках и мышечных волокнах происходит усиление обмена веществ. Это проявляется как в ряде биохимических изменений, происходящих в мембране и протоплазме клеток, так и в усилении их теплопродукции. Установлено, что при возбуждении происходят: усиление распада в клетках богатых энергией соединений — АТФ и креатинфосфата (КФ), усиление процессов распада и синтеза углеводов, белков и липидов, усиление окислительных процессов, приводящих в сочетании с гликолизом к резинтезу АТФ и КФ, синтез и разрушение ацетилхолина и норадреналина, других медиаторов, усиление синтеза РНК и белков. Все эти процессы более всего выражены в период восстановления состояния мембраны после ПД.
Все эти процессы более всего выражены в период восстановления состояния мембраны после ПД.
В нервах и мышцах каждая волна возбуждения сопровождается выделением двух порций тепла, из которых первая называется начальным, а вторая — запаздывающим теплом. Начальное теплообразование происходит в момент возбуждения и составляет незначительную часть всей теплопродукции (2-10%) при возбуждении. Предполагают, что это тепло связано с теми физико химическими процессами, которые развиваются в момент генерации ПД. Запаздывающее теплообразование происходит в течение более продолжительного времени, и длится многие минуты. Оно связано с теми химическими процессами, которые происходят в ткани вслед за волной возбуждения, и , по образному выражению Ухтомского, составляют «метаболический хвост кометы возбуждения».
Проведение возбуждения. Классификация нервных волокон. Как только в какой-либо точке нервного или мышечного волокна возникает ПД и этот участок приобретает отрицательный заряд, между возбужденными и соседними покоящимися участками волокна возникает электрический ток. В данном случае возбужденный участок мембраны действует на соседние участки как катод постоянного тока, вызывая их деполяризацию и генерируя локальный ответ. Если величина локального ответа превысит Ек мембраны, возникает ПД. В результате наружная поверхность мембраны заряжается отрицательно на новом участке. Таким способом волна возбуждения распространяется вдоль всего волокна со скоростью около 0,5-3 м/сек.
В данном случае возбужденный участок мембраны действует на соседние участки как катод постоянного тока, вызывая их деполяризацию и генерируя локальный ответ. Если величина локального ответа превысит Ек мембраны, возникает ПД. В результате наружная поверхность мембраны заряжается отрицательно на новом участке. Таким способом волна возбуждения распространяется вдоль всего волокна со скоростью около 0,5-3 м/сек.
Законы проведения возбуждения по нервам.
1. Закон физиологической непрерывности. Перерезка, перевязка, а также любое другое воздействие, нарушающее целость мембраны (физиологическую, а не только анатомическую), создают непроводимость. То же возникает при тепловых и химических воздействиях.
2. Закон двустороннего проведения. При нанесении раздражения на нервное волокно возбуждение распространяется по нему в обеих направлениях ( по поверхности мембраны — во все стороны) с одинаковой скоростью. Это доказывается опытом Бабухина и подобными ему.
Это доказывается опытом Бабухина и подобными ему.
3. Закон изолированного проведения. В нерве импульсы распространяются по каждому волокну изолированно, т.е. не переходят с одного волокна на другое. Это очень важно, так как обеспечивает точную адресовку импульса. Связано это с тем, что электрическое сопротивление миэлиновых и швановской оболочек, а также межклеточной жидкости значительно больше, чем сопротивление мембраны нервных волокон.
Механизмы и скорость проведения возбуждения в безмякотных и мякотных нервных волокнах различны. В безмякотных возбуждение распространяется непрерывно вдоль всей мембраны от одного возбужденного участка к другому, расположенному рядом, так, как мы уже обсуждали.
В миэлиновых волокнах возбуждение распространяется только скачкообразно, перепрыгивая через участки, покрытые миэлиновой оболочкой (сальтаторно). Потенциалы действия в этих волокнах возникают только в перехватах Ранвье. В состоянии покоя наружная поверхность возбудимой мембраны всех перехватов Ранвье заряжена положительно. В момент возбуждения поверхность первого перехвата становится отрицательно заряженной по отношению к соседнему второму перехвату. Это приводит к возникновению местного (локального) электротока, который идет через окружающую волокно межклеточную жидкость, мембрану и аксоплазму от перехвата 2 к 1. Выходящий через перехват 2 ток возбуждает его, вызывая перезарядку мембраны. Теперь этот участок может возбудить следующий и т.д.
В момент возбуждения поверхность первого перехвата становится отрицательно заряженной по отношению к соседнему второму перехвату. Это приводит к возникновению местного (локального) электротока, который идет через окружающую волокно межклеточную жидкость, мембрану и аксоплазму от перехвата 2 к 1. Выходящий через перехват 2 ток возбуждает его, вызывая перезарядку мембраны. Теперь этот участок может возбудить следующий и т.д.
Перепрыгивание ПД через межперехватный участок возможно потому, что амплитуда ПД в 5-6 раз больше порога, необходимого для возбуждения не только следующего, но и 3-5 перехватов. Поэтому микроповреждения волокна в межперехватных участках или не одном перехвате не прекращают работы нервного волокна до тех пор, пока регенеративный явления не захватят 3 и более лежащих рядом швановские клетки.
Время, необходимое для передачи возбуждения от одного перехвата к другому, одинаково у волокон различного диаметра, и составляют 0,07 мсек. Однако поскольку длина межперехватных участков различна и пропорциональна диаметру волокна, в миэлинизированных нервах скорость проведения нервных импульсов прямо пропорциональная их диаметру.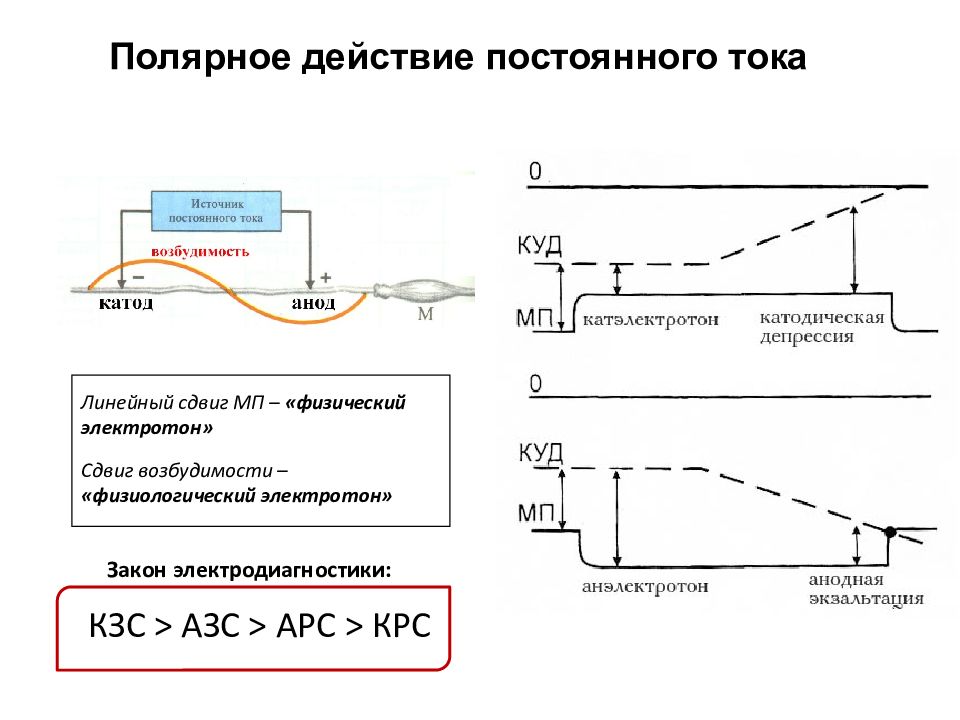
Классификация нервных волокон. Электрический ответ целого нерва является алгебраической суммой ПД отдельных его нервных волокон. Поэтому, с одной стороны, амплитуда электрических импульсов целого нерва зависит от силы раздражителя (с ее ростом вовлекаются все новые волокна), а во-вторых, суммарный потенциал действия нерва может быть расчленен на несколько отдельных колебаний, причиной чего является неодинаковая скорость проведения импульсов по разным волокнам, составляющим целый нерв.
В настоящее время нервные волокна по скорости проведения возбуждения, длительности различных фаз ПД и строении принято разделять на три основных типа.
Волокна типа А делятся на подгруппы (альфа, бета, гамма, дельта). Они покрыты миэлиновой оболочкой. Скорость проведения у них самая большая — 70-120 м/сек. Это — двигательные волокна, от моторных нейронов спинного мозга. Остальные волокна типа А — чувствительные.
Волокна типа В — миэлиновые, преимущественно преганглионарные.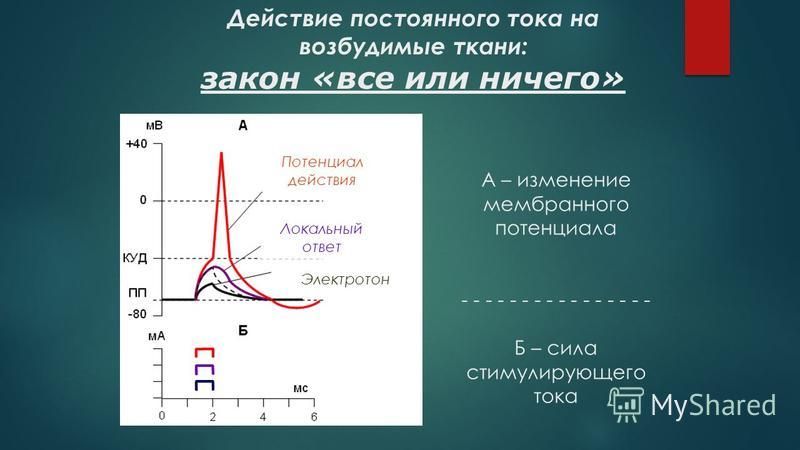 Скорость проведения — 3-18 м/сек.
Скорость проведения — 3-18 м/сек.
Волокна типа С — безмякотные, очень малого диаметра (2 мк). Скорость проведения не больше 3 м/сек. Это постганглионарные волокна симпатической нервной системы чаще всего.
Транскраниальная стимуляция постоянным током: протоколы и физиологические механизмы действия
Ziemann U, Paulus W, Nitsche MA, Pascual-Leone A, Byblow WD, Berardelli A, et al. Консенсус: протоколы пластичности моторной коры. Мозговой стимул. 2008; 1: 164–82.
Перекрёстная ссылка пабмед Google Scholar
Биндман Л., Липпольд О., Редферн Дж.В.Т. Действие кратковременных поляризующих токов на кору головного мозга крысы (1) при протекании тока и (2) при возникновении длительного последействия. Дж Физиол Лонд. 1964;172:369–82.
КАС ПабМед Центральный пабмед Google Scholar
 «>
«>Раш С., Дрисколл Д.А. Распределение тока в мозгу от поверхностных электродов. Анест Анальг. 1968; 47: 717–23.
Перекрёстная ссылка КАС пабмед Google Scholar
Даймонд А.М., Когер Р.В., Серафетинидес Э.А. Уровни внутримозгового тока у человека при терапии электросном. Биол психиатрия. 1975;10:101–4.
КАС пабмед Google Scholar
Пфуртшеллер Г. Спектральный анализ ЭЭГ: до, во время и после экстракраниальной стимуляции у человека. Электромед Биомед Тех. 1970; 15: 225–30. Статья на немецком языке.
Перекрёстная ссылка КАС пабмед Google Scholar
Костейн Р., Редферн Дж.В.Т., Липпольд OCJ. Контролируемое исследование терапевтического эффекта поляризации мозга при депрессивном заболевании. Бр Дж. Психиатрия. 1964;110:786–99.
Перекрёстная ссылка КАС пабмед Google Scholar
Липпольд OCJ, Redfearn JWT. Психические изменения, возникающие в результате прохождения малых постоянных токов через мозг человека. Бр Дж. Психиатрия. 1964; 1964: 768–72.
Перекрёстная ссылка Google Scholar
Redfearn JWT, Lippold OCJ, Costain R. Предварительный отчет о клинических эффектах поляризации мозга при некоторых психических расстройствах. Бр Дж. Психиатрия. 1964;110:773–85.
Перекрёстная ссылка КАС пабмед Google Scholar
Лолас Ф. Поляризация мозга: поведенческие и терапевтические эффекты. Биол психиатрия. 1977; 12:37–47.
КАС пабмед Google Scholar
Elbert T, Lutzenberger W, Rockstroh B, Birbaumer N.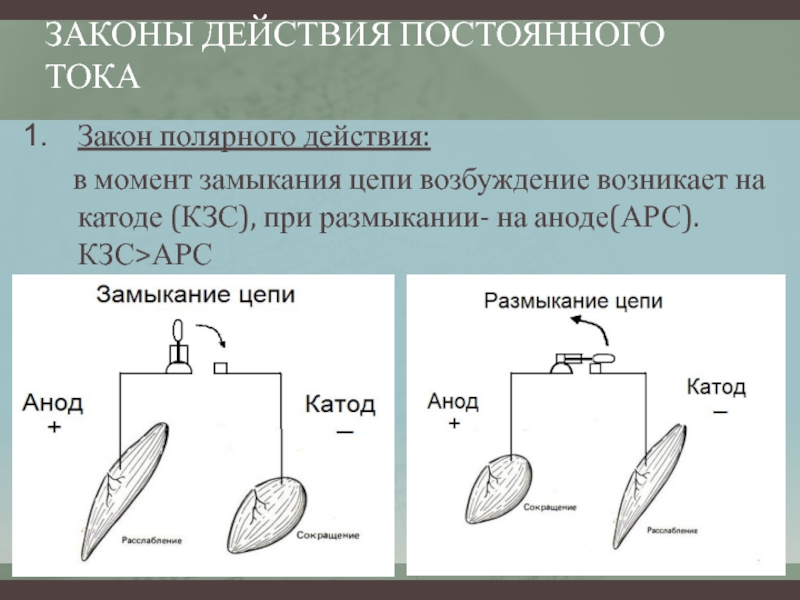 Влияние низкоуровневых транскортикальных постоянных токов на скорость реакции человека. Int J Neurosci. 1981;14:101–14.
Влияние низкоуровневых транскортикальных постоянных токов на скорость реакции человека. Int J Neurosci. 1981;14:101–14.
Перекрёстная ссылка КАС пабмед Google Scholar
Jaeger D, Elbert T, Lutzenberger W, Birbaumer N. Влияние внешних трансцефальных слабых постоянных токов на латерализацию в задачах реакции выбора. Дж. Психофизиол. 1987; 1: 127–33.
Google Scholar
Ницше М., Паулюс В. Изменения возбудимости, вызванные в моторной коре человека слабой транскраниальной стимуляцией постоянным током. Дж. Физиол. 2000; 527 (часть 3): 633–9..
Перекрёстная ссылка КАС ПабМед Центральный пабмед Google Scholar
 Неврология. 2001; 57: 1899–901.
Неврология. 2001; 57: 1899–901.Перекрёстная ссылка КАС пабмед Google Scholar
Ницше М.А., Коэн Л.Г., Вассерманн Э.М., Приори А., Ланг Н., Антал А. и другие. Транскраниальная стимуляция постоянным током: современное состояние 2008. Стимуляция мозга. 2008; 1: 206–23.
Перекрёстная ссылка пабмед Google Scholar
Ницше М.А., Паулюс В. Транскраниальная стимуляция постоянным током: обновление 2011. Restor Neurol Neurosci. 2011; 29: 463–92.
ПабМед Google Scholar
Dundas JE, Thickbroom GW, Mataglia FL. Восприятие комфорта при транскраниальной стимуляции постоянным током: влияние концентрации раствора NaCl на губчатые электроды. Клин Нейрофизиол. 2007; 118:1166–70.
Перекрёстная ссылка КАС пабмед Google Scholar
 «>
«>Ницше М., Ницше М., Кляйн С., Тергау Ф., Ротвелл Дж., Паулюс В. Уровень действия катодной поляризации постоянного тока, индуцированной ингибированием моторной коры человека. Клин Нейрофизиол. 2003; 114: 600–4.
Перекрёстная ссылка пабмед Google Scholar
Ницше М., Нихаус Л., Хоффманн К., Хенгст С., Либетанц Д., Паулюс В. и др. МРТ-исследование головного мозга человека, подвергнутого слабой стимуляции постоянным током лобной коры. Клин Нейрофизиол. 2004;115:2419–23.
Перекрёстная ссылка КАС пабмед Google Scholar
Айер М., Матту У., Графман Дж., Ломарев М., Сато С., Вассерманн Э. Безопасность и когнитивный эффект поляризации головного мозга лобного постоянного тока у здоровых людей. Неврология. 2005; 64: 872–5.
Перекрёстная ссылка КАС пабмед Google Scholar
 «>
«>Либетанц Д., Кох Р., Майенфельс С., Кониг Ф., Паулюс В., Ницше М.А. Пределы безопасности катодной транскраниальной стимуляции постоянным током у крыс. Клин Нейрофизиол. 2009 г.;120:1161–7.
Перекрёстная ссылка пабмед Google Scholar
Амбрус Г.Г., Антал А., Паулюс В. Сравнение кожного восприятия, вызванного электрической стимуляцией с использованием электродов прямоугольной и круглой формы. Клин Нейрофизиол. 2011; 122:803–7.
Перекрёстная ссылка пабмед Google Scholar
Амбрус Г.Г., Аль-Мойед Х., Чайеб Л., Сарп Л., Антал А., Паулюс В. Подход постепенного появления — короткая стимуляция — затухание к фиктивной tDCS — надежен при 1 мА для наивных и опытных субъектов, но не следователи. Мозговой стимул. 2012;5:499–504.
Перекрёстная ссылка пабмед Google Scholar
 «>
«>Paulus W. О трудностях отделения сетчатки от кортикального происхождения фосфенов при использовании транскраниальной стимуляции переменным током (tACS). Клин Нейрофизиол. 2010; 121:987–91.
Перекрёстная ссылка пабмед Google Scholar
Gandiga PC, Hummel FC, Cohen LG. Транскраниальная стимуляция постоянного тока (tDCS): инструмент для двойных слепых плацебо-контролируемых клинических исследований стимуляции мозга. Клин Нейрофизиол. 2006; 117: 845–50.
Перекрёстная ссылка пабмед Google Scholar
Miranda PC, Faria P, Hallett M. Что отношение вводимого тока к площади электрода говорит нам о плотности тока в мозге во время tDCS? Клин Нейрофизиол. 2009; 120:1183–1187.
Перекрёстная ссылка ПабМед Центральный пабмед Google Scholar
 «>
«>Фариа П., Халлетт М., Миранда П.С. Анализ методом конечных элементов влияния площади электродов и межэлектродного расстояния на пространственное распределение плотности тока в tDCS. Дж. Нейронная инженерия. 2011;8:066017.
Перекрёстная ссылка ПабМед Центральный пабмед Google Scholar
Пурпура DP, McMurtry JG. Изменения внутриклеточной активности и вызванных потенциалов при поляризации моторной коры. J Нейрофизиол. 1965; 28: 166–85.
КАС пабмед Google Scholar
Крейтцфельдт О.Д., Фромм Г.Х., Капп Х. Влияние транскортикальных постоянных токов на активность корковых нейронов. Опыт Нейрол. 1962;5:436–52.
Перекрёстная ссылка КАС пабмед Google Scholar
Кабаков А.Ю., Мюллер П.А., Паскуаль-Леоне А., Дженсен Ф. Е., Ротенберг А. Вклад ориентации аксонов в зависимую от пути модуляцию возбуждающей передачи путем стимуляции постоянным током в изолированном гиппокампе крысы. J Нейрофизиол. 2012; 107: 1881–9.
Е., Ротенберг А. Вклад ориентации аксонов в зависимую от пути модуляцию возбуждающей передачи путем стимуляции постоянным током в изолированном гиппокампе крысы. J Нейрофизиол. 2012; 107: 1881–9.
Перекрёстная ссылка ПабМед Центральный пабмед Google Scholar
Монте-Сильва К., Куо М.Ф., Хессенталер С., Фресноза С., Либетанц Д., Паулюс В. и др. Индукция поздней LTP-подобной пластичности в моторной коре человека повторной неинвазивной стимуляцией мозга. Мозговой стимул. 2012; 6: 424–32.
Перекрёстная ссылка пабмед Google Scholar
Аккорнеро Н., Ли Воти П., Ла Ричча М., Грегори Б. Модуляция зрительных вызванных потенциалов при поляризации коры постоянным током. Опыт Мозг Res. 2007; 178: 261–6.
Перекрёстная ссылка пабмед Google Scholar
 «>
«>Антал А., Терней Д., Порейс С., Паулюс В. На пути к разгадке связанных с задачей модуляций нейропластических изменений, вызванных в моторной коре человека. Евр Джей Нейроски. 2007; 26: 2687–91.
Перекрёстная ссылка пабмед Google Scholar
Рот Б.Дж. Механизмы электростимуляции возбудимой ткани. Crit Rev Biomed Eng. 1994;22:253–305.
КАС пабмед Google Scholar
Приори А., Берарделли А., Рона С., Аккорнеро Н., Манфреди М. Поляризация моторной коры человека через кожу головы. Нейроотчет. 1998; 9: 2257–60.
Перекрёстная ссылка КАС пабмед Google Scholar
Antal A, Kincses TZ, Nitsche MA, Bartfai O, Paulus W. Изменения возбудимости, вызванные транскраниальной стимуляцией постоянным током в первичной зрительной коре человека: прямые электрофизиологические данные. Invest Ophthalmol Vis Sci. 2004;45:702–7.
Invest Ophthalmol Vis Sci. 2004;45:702–7.
Перекрёстная ссылка пабмед Google Scholar
Молиадзе В., Антал А., Паулюс В. Последствия транскраниальной прямой и случайной шумовой стимуляции с экстрацефальными референтными электродами в зависимости от расстояния между электродами. Клин Нейрофизиол. 2010; 121:2165–71.
Перекрёстная ссылка пабмед Google Scholar
Монте-Сильва К., Куо М.-Ф., Либетанц Д., Паулюс В., Ницше М.А. Формирование оптимального интервала повторения катодной транскраниальной стимуляции постоянным током (tDCS). J Нейрофизиол. 2010; 103:1735–40.
Перекрёстная ссылка пабмед Google Scholar
Фрегни Ф., Хименес Р., Валье А.С., Феррейра М.Дж., Роша Р.Р., Наталле Л. и др. Рандомизированное, плацебо-контролируемое, подтверждающее принцип исследование транскраниальной стимуляции постоянным током для лечения боли при фибромиалгии. Ревмирующий артрит. 2006; 54:3988–98.
Ревмирующий артрит. 2006; 54:3988–98.
Перекрёстная ссылка пабмед Google Scholar
Loo CK, Alonzo A, Martin D, Mitchell PB, Galvez V, Sachdev P. Транскраниальная стимуляция постоянным током при депрессии: 3-недельное рандомизированное плацебо-контролируемое исследование. Бр Дж. Психиатрия. 2012; 200:52–9.
Перекрёстная ссылка пабмед Google Scholar
Рейс Дж., Шамбра Х.М., Коэн Л.Г., Бух Э.Р., Фрич Б., Заран Э. и др. Неинвазивная корковая стимуляция улучшает приобретение двигательных навыков в течение нескольких дней благодаря эффекту консолидации. Proc Natl Acad Sci U S A. 2009;106:1590–5.
Перекрёстная ссылка КАС ПабМед Центральный пабмед Google Scholar
Ланг Н., Зибнер Х., Уорд Н., Ли Л., Нитше М., Паулюс В. и др. Как транскраниальная стимуляция постоянным током первичной моторной коры изменяет активность региональных нейронов в мозге человека? Евр Джей Нейроски. 2005;22:495–504.
и др. Как транскраниальная стимуляция постоянным током первичной моторной коры изменяет активность региональных нейронов в мозге человека? Евр Джей Нейроски. 2005;22:495–504.
Перекрёстная ссылка ПабМед Центральный пабмед Google Scholar
Ницше М.А., Доемкес С., Каракосе Т., Антал А., Либетанц Д., Ланг Н. и другие. Формирование эффектов транскраниальной стимуляции постоянным током моторной коры человека. J Нейрофизиол. 2007;97:3109–17.
Перекрёстная ссылка КАС пабмед Google Scholar
Датта А., Бансал В., Диаз Дж., Патель Дж., Реато Д., Биксон М. Точная модель головы для транскраниальной стимуляции постоянным током: улучшенная пространственная фокусировка с использованием кольцевого электрода по сравнению с обычной прямоугольной подкладкой. Мозговой стимул. 2009 г.;2:201–7. 207.e201.
Перекрёстная ссылка ПабМед Центральный пабмед Google Scholar
 «>
«>Cogiamanian F, Marceglia S, Ardolino G, Barbieri S, Priori A. Повышение выносливости к изометрической силе после транскраниальной стимуляции постоянным током двигательных областей коры головного мозга человека. Евр Джей Нейроски. 2007; 26: 242–9.
Перекрёстная ссылка КАС пабмед Google Scholar
Borckardt JJ, Bikson M, Frohman H, Reeves ST, Datta A, Bansal V, et al. Пилотное исследование переносимости и влияния транскраниальной стимуляции постоянным током высокой четкости (HD-tDCS) на восприятие боли. Джей Пейн. 2012;13:112–20.
Перекрёстная ссылка пабмед Google Scholar
Дмоховски Дж.П., Датта А., Биксон М., Су Ю., Парра Л.С. Оптимизированная мультиэлектродная стимуляция увеличивает фокус и интенсивность воздействия на цель. Дж. Нейронная инженерия. 2011;8:046011.
Перекрёстная ссылка пабмед Google Scholar
 «>
«>Hallett M. Транскраниальная магнитная стимуляция: введение. Нейрон. 2007; 55: 187–99.
Перекрёстная ссылка КАС пабмед Google Scholar
Нитше М., Сибер А., Фромманн К., Кляйн С., Рочфорд С., Нитше М. и др. Модулирующие параметры возбудимости во время и после транскраниальной стимуляции постоянным током моторной коры человека. Дж. Физиол. 2005; 568 (часть 1): 291–303.
Перекрёстная ссылка КАС ПабМед Центральный пабмед Google Scholar
Ницше М., Либетанц Д., Шлиттерлау А., Хеншке У., Фрике К., Фромманн К. и др. ГАМКергическая модуляция сдвигов возбудимости моторной коры, вызванных стимуляцией постоянного тока, у людей. Евр Джей Нейроски. 2004;19: 2720–6.
Перекрёстная ссылка пабмед Google Scholar
Антал А. , Паулюс В. Исследование нейропластических изменений в головном мозге человека, вызванных методами транскраниальной прямой (tDCS) и переменного тока (tACS) стимуляции. Клин ЭЭГ Neurosci. 2012;43:175.
, Паулюс В. Исследование нейропластических изменений в головном мозге человека, вызванных методами транскраниальной прямой (tDCS) и переменного тока (tACS) стимуляции. Клин ЭЭГ Neurosci. 2012;43:175.
Перекрёстная ссылка пабмед Google Scholar
Борос К., Порейс К., Мюнхау А., Паулюс В., Ницше М.А. Премоторная транскраниальная стимуляция постоянным током (tDCS) влияет на первичную моторную возбудимость человека. Евр Джей Нейроски. 2008;27:1292–300.
Перекрёстная ссылка пабмед Google Scholar
Нитше М., Фрике К., Хеншке У., Шлиттерлау А., Либетанц Д., Ланг Н. и др. Фармакологическая модуляция сдвигов возбудимости коры, вызванных транскраниальной стимуляцией постоянным током у человека. Дж. Физиол. 2003; 553: 293–301.
Перекрёстная ссылка КАС ПабМед Центральный пабмед Google Scholar
 «>
«>Ницше М., Яусси В., Либетанц Д., Ланг Н., Тергау Ф., Паулюс В. Консолидация моторной кортикальной нейропластичности человека с помощью D-циклосерина. Нейропсихофармакология. 2004;29: 1573–158.
Перекрёстная ссылка КАС пабмед Google Scholar
Ислам Н., Афтабуддин М., Мориваки А., Хаттори Ю., Хори Ю. Повышение уровня кальция после анодной поляризации в мозге крысы. Мозг Res. 1995; 684: 206–8.
Перекрёстная ссылка КАС пабмед Google Scholar
Stagg CJ, Best JG, Stephenson MC, O’Shea J, Wylezinska M, Kincses ZT и др. Чувствительная к полярности модуляция корковых нейротрансмиттеров при транскраниальной стимуляции. Дж. Нейроски. 2009 г.;29:5202–6.
Перекрёстная ссылка КАС пабмед Google Scholar
Рейманн К. Г., Фрей Ю.Ю. Позднее поддержание LTP гиппокампа: требования, фазы, «синаптическая маркировка», «поздняя ассоциативность» и последствия. Нейрофармакология. 2007; 52:24–40.
Г., Фрей Ю.Ю. Позднее поддержание LTP гиппокампа: требования, фазы, «синаптическая маркировка», «поздняя ассоциативность» и последствия. Нейрофармакология. 2007; 52:24–40.
Перекрёстная ссылка КАС пабмед Google Scholar
Ницше М.А., Мюллер-Дальхаус Ф., Паулюс В., Циманн У. Фармакология нейропластичности, вызванной неинвазивной стимуляцией мозга: создание моделей для клинического применения препаратов, действующих на ЦНС. Дж. Физиол. 2012;590: 4641–62.
Перекрёстная ссылка КАС ПабМед Центральный пабмед Google Scholar
Ницше М., Гранди Дж., Либетанц Д., Ланг Н., Тергау Ф., Паулюс В. Катехоламинергическая консолидация моторной корковой нейропластичности у людей. Кора головного мозга. 2004; 14:1240–5.
Перекрёстная ссылка пабмед Google Scholar
 «>
«>Нитше М., Лампе С., Антал А., Либетанц Д., Ланг Н., Тергау Ф. и др. Дофаминергическая модуляция длительных изменений возбудимости коры, индуцированных постоянным током, в моторной коре человека. Евр Джей Нейроски. 2006; 23:1651–7.
Перекрёстная ссылка пабмед Google Scholar
Куо М-Ф, Паулюс В., Ницше М.А. Повышение очагово-индуцированной пластичности мозга дофамином. Кора головного мозга. 2008; 18: 648–51.
Перекрёстная ссылка пабмед Google Scholar
Монте-Сильва К., Либетанц Д., Грандей Дж., Паулюс В., Ницше М.А. Нелинейный эффект леводопы в зависимости от дозировки на пластичность моторной коры человека. Дж Физиол Лонд. 2010; 588:3415–24.
Перекрёстная ссылка КАС ПабМед Центральный пабмед Google Scholar
Монте-Сильва К. , Куо М.Ф., Тиругнансамбандам Н., Либетанц Д., Паулюс В., Ницше М.А. Дозозависимый перевернутый U-образный эффект активации дофаминовых (D2-подобных) рецепторов на фокальную и нефокальную пластичность у людей. Дж. Нейроски. 2009; 29: 6124–31.
, Куо М.Ф., Тиругнансамбандам Н., Либетанц Д., Паулюс В., Ницше М.А. Дозозависимый перевернутый U-образный эффект активации дофаминовых (D2-подобных) рецепторов на фокальную и нефокальную пластичность у людей. Дж. Нейроски. 2009; 29: 6124–31.
Перекрёстная ссылка КАС пабмед Google Scholar
Ницше М.А., Куо М.Ф., Грош Дж., Бергнер С., Монте-Сильва К., Паулюс В. Влияние D1-рецептора на нейропластичность у людей. Дж. Нейроски. 2009; 29: 2648–53.
Перекрёстная ссылка КАС пабмед Google Scholar
Ницше М.А., Куо М.Ф., Карраш Р., Вехтер Б., Либетанц Д., Паулюс В. Серотонин влияет на транскраниальную нейропластичность, индуцированную постоянным током, у людей. Биол психиатрия. 2009; 66: 503–8.
Перекрёстная ссылка КАС пабмед Google Scholar
 «>
«>Куо М.-Ф., Грош Дж., Фрегни Ф., Паулюс В., Ницше М.А. Фокусирующее влияние ацетилхолина на нейропластичность моторной коры человека. Дж. Нейроски. 2007; 27:14442–7.
Перекрёстная ссылка КАС пабмед Google Scholar
Тиругнансамбандам Н., Гранди Дж., Адам К., Дрис А., Сквирба А.С., Ланг Н. и др. Никотинергическое влияние на очаговую и неочаговую нейропластичность, вызванную неинвазивной стимуляцией мозга у некурящих людей. Нейропсихофармакология. 2011;36:879–86.
Перекрёстная ссылка КАС ПабМед Центральный пабмед Google Scholar
Мацунага К., Ницше М.А., Цудзи С., Ротвелл Дж.К. Влияние транскраниальной стимуляции сенсомоторной коры постоянного тока на соматосенсорные вызванные потенциалы у человека. Клин Нейрофизиол. 2004; 115:456–60.
Перекрёстная ссылка пабмед Google Scholar
 «>
«>Dieckhofer A, Waberski TD, Nitsche M, Paulus W, Buchner H, Gobbele R. Транскраниальная стимуляция постоянным током, применяемая к соматосенсорной коре: дифференциальное воздействие на низкочастотные и высокочастотные SEP. Клин Нейрофизиол. 2006; 117: 2221–7.
Перекрёстная ссылка пабмед Google Scholar
Zaehle T, Beretta M, Jancke L, Herrmann CS, Sandmann P. Изменения возбудимости, вызванные транскраниальной стимуляцией постоянным током в слуховой коре человека: прямые электрофизиологические данные. Опыт Мозг Res. 2011; 215:135–40.
Перекрёстная ссылка пабмед Google Scholar
Kirimoto H, Ogata K, Onishi H, Oyama M, Goto Y, Tobimatsu S. Транскраниальная стимуляция постоянным током моторной ассоциативной коры вызывает пластические изменения в ипсилатеральной первичной моторной и соматосенсорной коре. Клин Нейрофизиол. 2011; 122:777–83.
2011; 122:777–83.
Перекрёстная ссылка пабмед Google Scholar
Feurra M, Bianco G, Polizzotto NR, Innocenti I, Rossi A, Rossi S. Корково-корковая связь между правой теменной и двусторонней первичной моторной корой во время воображаемых и наблюдаемых действий: комбинированное исследование TMS/tDCS. Передние нейронные цепи. 2011;5:10.
Перекрёстная ссылка ПабМед Центральный пабмед Google Scholar
Полания Р., Паулюс В., Антал А., Ницше М.А. Введение теории графов для отслеживания нейропластических изменений в мозге человека в состоянии покоя: исследование транскраниальной стимуляции постоянным током. Нейроизображение. 2011;54:2287–96.
Перекрёстная ссылка пабмед Google Scholar
Stagg CJ, O’Shea J, Kincses ZT, Woolrich M, Matthews PM, Johansen-Berg H. Модуляция связанной с движением активации коры с помощью транскраниальной стимуляции постоянным током. Евр Джей Нейроски. 2009 г.;30:1412–23.
Модуляция связанной с движением активации коры с помощью транскраниальной стимуляции постоянным током. Евр Джей Нейроски. 2009 г.;30:1412–23.
Перекрёстная ссылка КАС пабмед Google Scholar
Полания Р., Ницше М.А., Паулюс В. Модулирование паттернов функциональной связи и топологической функциональной организации человеческого мозга с помощью транскраниальной стимуляции постоянным током. Hum Brain Map. 2011;32:1236–49.
Перекрёстная ссылка пабмед Google Scholar
Полания Р., Паулюс В., Ницше М.А. Модулирование корково-стриарной и таламо-кортикальной функциональной связи с помощью транскраниальной стимуляции постоянным током. Hum Brain Map. 2011;32(8):1236–49..
Перекрёстная ссылка пабмед Google Scholar
Полания Р. , Паулюс В., Ницше М.А. Реорганизация внутренней функциональной архитектуры первичной моторной коры человека во время отдыха с помощью неинвазивной стимуляции коры. ПЛОС ОДИН. 2012;7:e30971.
, Паулюс В., Ницше М.А. Реорганизация внутренней функциональной архитектуры первичной моторной коры человека во время отдыха с помощью неинвазивной стимуляции коры. ПЛОС ОДИН. 2012;7:e30971.
Перекрёстная ссылка КАС ПабМед Центральный пабмед Google Scholar
Кизер Д., Майндл Т., Бор Дж., Палм Ю., Погарелл О., Мулерт С. и др. Префронтальная транскраниальная стимуляция постоянным током изменяет связность сетей в состоянии покоя во время фМРТ. Дж. Нейроски. 2011; 31:15284–93.
Перекрёстная ссылка КАС пабмед Google Scholar
Пена-Гомез С., Сала-Лонч Р., Юнке С., Клементе И.С., Видаль Д., Баргалло Н. и др. Модуляция крупномасштабных сетей мозга путем транскраниальной стимуляции постоянным током, подтвержденная функциональной МРТ в состоянии покоя. Мозговой стимул. 2012;5:252–63.
Перекрёстная ссылка ПабМед Центральный пабмед Google Scholar
 «>
«>Либетанц Д., Ницше М.А., Тергау Ф., Паулюс В. Фармакологический подход к механизмам последействия возбудимости моторной коры человека, вызванного транскраниальной стимуляцией постоянным током. Мозг. 2002; 125:2238–47.
Перекрёстная ссылка пабмед Google Scholar
Куо Х.И., Биксон М., Датта А., Минхас П., Паулюс В., Куо М.Ф. и др. Сравнение кортикальной пластичности, вызванной обычной и кольцевой tDCS 4×1 высокого разрешения: нейрофизиологическое исследование. Мозговой стимул. 2012; 6: 644–8.
Перекрёстная ссылка пабмед Google Scholar
Что такое транскраниальная стимуляция постоянным током (tDCS)?
Основы транскраниальной стимуляции постоянным током (tDCS):
Транскраниальная стимуляция постоянным током (tDCS) — это портативный, носимый метод стимуляции мозга, который подает слабый электрический ток на кожу головы. Обычно применяется фиксированный ток от 1 до 2 мА 1 . tDCS работает, подавая положительный (анодный) или отрицательный (катодный) ток через электроды на область. tDCS — это метод нейромодуляции, который вызывает немедленные и длительные изменения в функции мозга. Положение анодного и катодного электродов на голове используется для определения того, как ток течет к определенным областям мозга. Ток, подаваемый tDCS, НЕ достаточно силен, чтобы вызвать потенциал действия в нейроне; вместо этого его «подпорог» изменяет структуру уже активных нейронов. Думайте о мозге как об активном человеке, пытающемся что-то сделать или чему-то научиться, и tDCS, подстегивающем эту постоянную активность. На клеточном уровне 2 , tDCS изменяет возбуждение нейронов и, усиливая синаптическую передачу между нейронами, увеличивает синаптическую пластичность 3 , которая, в свою очередь, является клеточной основой обучения. tDCS часто сочетают с обучением. Тренировка сама по себе производит обучение (синаптическая пластичность), а одновременная tDCS усиливает эти эффекты (повышает синаптическую пластичность).
Обычно применяется фиксированный ток от 1 до 2 мА 1 . tDCS работает, подавая положительный (анодный) или отрицательный (катодный) ток через электроды на область. tDCS — это метод нейромодуляции, который вызывает немедленные и длительные изменения в функции мозга. Положение анодного и катодного электродов на голове используется для определения того, как ток течет к определенным областям мозга. Ток, подаваемый tDCS, НЕ достаточно силен, чтобы вызвать потенциал действия в нейроне; вместо этого его «подпорог» изменяет структуру уже активных нейронов. Думайте о мозге как об активном человеке, пытающемся что-то сделать или чему-то научиться, и tDCS, подстегивающем эту постоянную активность. На клеточном уровне 2 , tDCS изменяет возбуждение нейронов и, усиливая синаптическую передачу между нейронами, увеличивает синаптическую пластичность 3 , которая, в свою очередь, является клеточной основой обучения. tDCS часто сочетают с обучением. Тренировка сама по себе производит обучение (синаптическая пластичность), а одновременная tDCS усиливает эти эффекты (повышает синаптическую пластичность). В настоящее время изучаются некоторые клинические применения tDCS: депрессия, шизофрения, афазия, зависимость 4 , эпилепсия, хроническая боль (мигрень, фибромиалгия), внимание и двигательная реабилитация. tDCS также используется для немедицинских оздоровительных приложений, например, для ускоренного обучения 5 , концентрация, расслабление и медитация. 6
В настоящее время изучаются некоторые клинические применения tDCS: депрессия, шизофрения, афазия, зависимость 4 , эпилепсия, хроническая боль (мигрень, фибромиалгия), внимание и двигательная реабилитация. tDCS также используется для немедицинских оздоровительных приложений, например, для ускоренного обучения 5 , концентрация, расслабление и медитация. 6
Как выглядит устройство для транскраниальной стимуляции постоянным током (tDCS)?
Устройства tDCS представляют собой небольшие устройства с батарейным питанием. Обычно имеется панель управления, которая позволяет программировать устройство (устанавливать продолжительность и интенсивность стимуляции). Электроды надеваются на голову и удерживаются головным убором — обычно эластичным ремешком. Кабель соединяет каждый электрод со стимулятором. При включении стимулятора ток идет от устройства к электроду, а затем через мозг. Стимуляторы профессионального уровня имеют множество функций, которые помогают обеспечить переносимость и надежность стимуляции. Это включает в себя измеритель импеданса и измеритель тока.
Это включает в себя измеритель импеданса и измеритель тока.
Каковы побочные эффекты транскраниальной стимуляции постоянным током (tDCS)?
Исследования побочных эффектов tDCS продолжаются, но пока установленные побочные эффекты незначительны 7 и ограничиваются расположением электрода. Они включают временное покраснение кожи, зуд и покалывание. Другие предполагаемые побочные эффекты tDCS включают головную боль, тошноту и головокружение. Следует отметить, что эти последние три побочных эффекта, как было показано, проявляются почти с той же частотой, что и ложная стимуляция (фальшивая стимуляция) 8 При неправильном применении tDCS могут возникать другие побочные эффекты, такие как фосфен, представляющий собой временную неопасную вспышку света. Это может произойти, если электроды расположены слишком близко к глазу. Кроме того, неправильное введение tDCS может вызвать стандартные ожоги кожи. Нет никаких научных доказательств, демонстрирующих стойкие травмы или необратимые побочные эффекты от tDCS. Тем не менее, следует отметить, что все данные о переносимости и безопасности tDCS получены в результате контролируемых испытаний на людях с использованием специализированного оборудования и строго контролируемых протоколов (например, ограничение текущей продолжительности, количество сеансов).
Тем не менее, следует отметить, что все данные о переносимости и безопасности tDCS получены в результате контролируемых испытаний на людях с использованием специализированного оборудования и строго контролируемых протоколов (например, ограничение текущей продолжительности, количество сеансов).
На что похожа транскраниальная стимуляция постоянным током (tDCS)?
Во время tDCS большинство людей ощущают легкое покалывание, покалывание, зуд или тепло. Эти ощущения безболезненны и проходят при прекращении стимуляции. Однако «переносимость» tDCS зависит от качества принадлежностей, процедур настройки и использования интенсивности (несколько мА) и продолжительности (десятки минут), соответствующих стандартам tDCS. 23
Чего мы не знаем о транскраниальной стимуляции постоянным током (tDCS)?
В то время как остаются вопросы о лучших приложениях для tDCS, существуют десятилетия исследований, обозначающих его причастный механизм. Недавняя работа предполагает, что активация глии и изменение внутриклеточных концентраций цАМФ и кальция в значительной степени способствуют эффектам tDCS. временная потенциация (ДП) и длительная депрессия (ДД).
временная потенциация (ДП) и длительная депрессия (ДД).
Где можно пройти транскраниальную стимуляцию постоянным током (tDCS)?
В США tDCS имеет регулирующий статус «исследовательского» 10 . Это не указывает на эффективность; это означает, что FDA не выдало заключение. Как правило, FDA не дает заключения до тех пор, пока компании не проявят интерес к маркетингу устройства. В Соединенных Штатах компаниям не разрешается продавать tDCS по клиническим показаниям, таким как «лечение депрессии» или «лечение эпилепсии». Врачам в Соединенных Штатах разрешено проводить лечение «не по прямому назначению», то есть лечение, которое не одобрено FDA для данного показания. Исследовательским центрам по всему миру разрешено тестировать tDCS в контролируемых клинических испытаниях. В таких испытаниях каждый субъект должен подписать лист информированного согласия. Вы можете найти список испытаний tDCS здесь: Clinicaltrials.gov. В ЕС tDCS одобрен для лечения боли и депрессии. Вы можете использовать бесплатный инструмент ниже, чтобы найти врачей и клиники, которые предоставляют лечение на основе tDCS
Вы можете использовать бесплатный инструмент ниже, чтобы найти врачей и клиники, которые предоставляют лечение на основе tDCS
Одобрена ли tDCS FDA?
tDCS в настоящее время не одобрен FDA. Это означает, что FDA США не оценивало и не одобряло «маркетинговую» заявку от компании. Это не означает, что FDA США приняло официальное решение об эффективности или безопасности tDCS для индукции любой спецификации, такой как депрессия или боль. В США tDCS для медицинского применения считается «исследовательским». FDA не читает клинические испытания и принимает решения на основе литературы, FDA США отвечает только на «маркетинговые» запросы, сделанные конкретными компаниями. FDA обычно не регулирует немедицинское использование устройств, включая использование для «здоровья». цели 11 . В этом смысле важно отметить, что tDCS широко рассматривается исследователями и экспертами как заболевание с низким уровнем риска. Фактически, FDA предоставило письма «513g» нескольким компаниям, прямо разрешая им продавать tDCS для конкретных немедицинских целей.
FDA также не регулирует медицинскую практику, то есть не регулирует работу врачей. По этой причине многие врачи назначают лечение «не по прямому назначению» — то, что, по мнению врачей, работает, но не имеет «маркетингового» ярлыка от FDA для компании. tDCS одобрен для лечения в большинстве стран мира 12 , включая Европейский Союз 13 , Израиль и Сингапур. Таким образом, хотя tDCS в настоящее время не одобрена FDA, это не означает, что tDCS нельзя тестировать на законных основаниях или использовать в определенных контекстах.
Фактически, FDA предоставило письма «513g» нескольким компаниям, прямо разрешая им продавать tDCS для конкретных немедицинских целей.
FDA также не регулирует медицинскую практику, то есть не регулирует работу врачей. По этой причине многие врачи назначают лечение «не по прямому назначению» — то, что, по мнению врачей, работает, но не имеет «маркетингового» ярлыка от FDA для компании. tDCS одобрен для лечения в большинстве стран мира 12 , включая Европейский Союз 13 , Израиль и Сингапур. Таким образом, хотя tDCS в настоящее время не одобрена FDA, это не означает, что tDCS нельзя тестировать на законных основаниях или использовать в определенных контекстах.
Сколько стоит tDCS?
Стоимость устройств tDCS может варьироваться от примерно 100 долларов США за базовые «потребительские» устройства tDCS до тысяч долларов за системы tDCS исследовательского уровня. Существует широкий спектр функций и возможностей в различных
устройства.
Может ли tDCS лечить тревогу?
Эффекты tDCS по снижению тревожности были зарегистрированы в нескольких клинических испытаниях 14 , проведенных ведущим медицинским центром. В США tDCS не одобрен для лечения медицинской тревоги. Методы, связанные с tDCS, такие как транскраниальная стимуляция переменным током (tACS), показали многообещающие результаты в клинических испытаниях 15 . Другой родственный метод, стимуляция краниальной электротерапии (CES), одобрен FDA для тревоги 16 .
Может ли tDCS лечить депрессию?
Несколько клинических испытаний показали, что tDCS может лечить депрессию 17 . tDCS также имеет гораздо меньше побочных эффектов, чем лекарства 18 . В США tDCS не одобрен для лечения депрессии. В большинстве стран мира, в том числе в Европе, tDCS одобрен для лечения депрессии 19 .
Каковы преимущества tDCS?
tDCS используется для многих различных приложений, которые включают изменение мозга, чтобы влиять на то, как люди думают или чувствуют 20 . tDCS часто сочетается с какой-либо другой формой активности или обучения с целью tDCS повысить эту конкретную мозговую активность. Было показано, что tDCS заставляет людей учиться быстрее 21 . Например, tDCS может улучшить осознанность (электронная медитация). Люди также заинтересованы в tDCS для увеличения 22 «рабочей памяти».
tDCS часто сочетается с какой-либо другой формой активности или обучения с целью tDCS повысить эту конкретную мозговую активность. Было показано, что tDCS заставляет людей учиться быстрее 21 . Например, tDCS может улучшить осознанность (электронная медитация). Люди также заинтересованы в tDCS для увеличения 22 «рабочей памяти».
Поделиться |
Содержимое не предназначено для замены профессиональной медицинской консультации, диагностики или лечения. Всегда обращайтесь за советом к своему врачу или другому квалифицированному поставщику медицинских услуг по любым вопросам, которые могут у вас возникнуть относительно состояния здоровья.
1. Вудс А.Дж., Антал А., Биксон М., Богджио П.С., Брунони А.Р., Сельник П.… Ницше М.А. Техническое руководство по tDCS и связанным с ними инструментам неинвазивной стимуляции мозга. Клиническая нейрофизиология. 2016;127(2):1031–1048.
2. Выступление доктора Майкла Ницше о физиологических основах tDCS на YouTube. 3. Кронберг Г., Рахман А., Шарма М., Биксон М. и Парра Л., 2020. Стимуляция постоянным током повышает пластичность Хебба в лабораторных условиях. Стимуляция мозга
4. Эхтиари Х., Таваколи Х., Аддолорато Г., Баекен С., Бончи А., Кампанелла С., Каштело-Бранко Л., Шале-Буджу Г., Кларк В.П., Клаус Э., Дэннон П.Н., Дель Феличе А., Ден Уйл Т., Дайана М., ди Джаннантонио М., Федота Дж. Р., Фитцджеральд П., Галлимберти Л., Гралль-Броннек М., Херреманс СК, Херрманн М.Дж., Джамиль А., Хедр Э., Куимцидис С., Козак К., Крупицкий Э., Ламм С., Лехнер В.В., Мадео Г., Мальмир Н. , Мартинотти Г., Макдональд В., Монтемитро К., Накамура-Паласиос Э.М., Насехи М., Ноэль Х, Носратабади М., Паулюс М., Петторрусо М., Прадхан Б., Прахарадж С.К., Рафферти Х., Салем Г., Салмерон Б.Дж., Соважет А., Шлютер Р.С. , Сержиу К., Шахбабаи А., Шеффер К., Спаньоло П.А., Стил В.Р., Юань Т-Ф, ван Донген Дж., Ван Ваес В., Венкатасубраманян Г., ВердехоГарсия А., Вервир И., Уэлш Дж., Уэсли М.Дж., Виткевиц К., Явари Ф., Зарриндаст М.
3. Кронберг Г., Рахман А., Шарма М., Биксон М. и Парра Л., 2020. Стимуляция постоянным током повышает пластичность Хебба в лабораторных условиях. Стимуляция мозга
4. Эхтиари Х., Таваколи Х., Аддолорато Г., Баекен С., Бончи А., Кампанелла С., Каштело-Бранко Л., Шале-Буджу Г., Кларк В.П., Клаус Э., Дэннон П.Н., Дель Феличе А., Ден Уйл Т., Дайана М., ди Джаннантонио М., Федота Дж. Р., Фитцджеральд П., Галлимберти Л., Гралль-Броннек М., Херреманс СК, Херрманн М.Дж., Джамиль А., Хедр Э., Куимцидис С., Козак К., Крупицкий Э., Ламм С., Лехнер В.В., Мадео Г., Мальмир Н. , Мартинотти Г., Макдональд В., Монтемитро К., Накамура-Паласиос Э.М., Насехи М., Ноэль Х, Носратабади М., Паулюс М., Петторрусо М., Прадхан Б., Прахарадж С.К., Рафферти Х., Салем Г., Салмерон Б.Дж., Соважет А., Шлютер Р.С. , Сержиу К., Шахбабаи А., Шеффер К., Спаньоло П.А., Стил В.Р., Юань Т-Ф, ван Донген Дж., Ван Ваес В., Венкатасубраманян Г., ВердехоГарсия А., Вервир И., Уэлш Дж., Уэсли М.Дж., Виткевиц К., Явари Ф., Зарриндаст М. Р. , Zawertailo L, Zhang X, Cha YH, George TP, Frohlich F, Goudriaan AE, Fecteau S, Daughters SB, Stein EA, Fregni F, Nitsche MA, Zangen A, Bikson M, Hanlon CA (2019). Транскраниальная электрическая и магнитная стимуляция (tES и TMS) для лечения зависимости: согласованный документ о нынешнем состоянии науки и пути вперед. Нейронаука и биоповеденческие обзоры. 2019. 104: 118-140
5. Коффман Б., Трамбо М., Флорес Р., Гарсия К., ван дер Мерве А., Вассерманн Э., Вайзенд М. и Кларк В., 2020. Влияние Tdcs на Производительность и обучение обнаружению целей: взаимодействие с характеристиками стимула и экспериментальным планом.
6. Электронная медитация: новый инструмент для древней техники на сайте Medical X Press Медицинского университета Южной Каролины.
7. Биксон М., Гроссман П., Томас С., Занноу А.Л., Цзян Дж., Аднан Т. и др. (2016). Безопасность транскраниальной стимуляции постоянным током: обновление, основанное на фактических данных, 2016 г. Стимуляция мозга. 9641–661. 10.1016/j.brs.
Р. , Zawertailo L, Zhang X, Cha YH, George TP, Frohlich F, Goudriaan AE, Fecteau S, Daughters SB, Stein EA, Fregni F, Nitsche MA, Zangen A, Bikson M, Hanlon CA (2019). Транскраниальная электрическая и магнитная стимуляция (tES и TMS) для лечения зависимости: согласованный документ о нынешнем состоянии науки и пути вперед. Нейронаука и биоповеденческие обзоры. 2019. 104: 118-140
5. Коффман Б., Трамбо М., Флорес Р., Гарсия К., ван дер Мерве А., Вассерманн Э., Вайзенд М. и Кларк В., 2020. Влияние Tdcs на Производительность и обучение обнаружению целей: взаимодействие с характеристиками стимула и экспериментальным планом.
6. Электронная медитация: новый инструмент для древней техники на сайте Medical X Press Медицинского университета Южной Каролины.
7. Биксон М., Гроссман П., Томас С., Занноу А.Л., Цзян Дж., Аднан Т. и др. (2016). Безопасность транскраниальной стимуляции постоянным током: обновление, основанное на фактических данных, 2016 г. Стимуляция мозга. 9641–661. 10.1016/j.brs. 2017.07.001
8. Брунони А.Р., Амадера Дж., Бербель Б., Волц М.С., Риццерио Б.Г. и Фрегни Ф. (2011). Систематический обзор отчетов и оценки побочных эффектов, связанных с транскраниальной стимуляцией постоянным током. Международный журнал нейропсихофармакологии, 14(8), 1133-1145.
9. Монаи Х., Окура М., Танака М., Оэ Ю., Конно А., Хираи Х., … и Хирасе Х. (2016). Визуализация кальция показывает участие глии в пластичности, индуцированной транскраниальной стимуляцией постоянным током, в мозге мышей. Связи с природой, 7.
10. Использование продаваемых лекарств, биологических препаратов и медицинских устройств «не по прямому назначению» и исследовательское использование — Руководство для институциональных наблюдательных советов и клинических исследователей fda.gov
11. Общее оздоровление: политика в отношении устройств с низким уровнем риска — руководство для промышленности и персонала Управления по санитарному надзору за качеством пищевых продуктов и медикаментов fda.ggOyR0iXCbMQv3Xipma34MD
12.
2017.07.001
8. Брунони А.Р., Амадера Дж., Бербель Б., Волц М.С., Риццерио Б.Г. и Фрегни Ф. (2011). Систематический обзор отчетов и оценки побочных эффектов, связанных с транскраниальной стимуляцией постоянным током. Международный журнал нейропсихофармакологии, 14(8), 1133-1145.
9. Монаи Х., Окура М., Танака М., Оэ Ю., Конно А., Хираи Х., … и Хирасе Х. (2016). Визуализация кальция показывает участие глии в пластичности, индуцированной транскраниальной стимуляцией постоянным током, в мозге мышей. Связи с природой, 7.
10. Использование продаваемых лекарств, биологических препаратов и медицинских устройств «не по прямому назначению» и исследовательское использование — Руководство для институциональных наблюдательных советов и клинических исследователей fda.gov
11. Общее оздоровление: политика в отношении устройств с низким уровнем риска — руководство для промышленности и персонала Управления по санитарному надзору за качеством пищевых продуктов и медикаментов fda.ggOyR0iXCbMQv3Xipma34MD
12. Френьи Ф., Ницше М.А., Лоо К.К., Брунони А.Р., Маранголо П., Лейте Дж., Карвалью С., Болоньини Н., Каумо В., Пайк Н.Дж., Симис М., Уеда К., Эхтиари Х., Луу П., Такер Д.М., Тайлер В.Дж., Brunelin J, Datta A, Juan CH, Venkatasubramanian G, Boggio PM, Bikson M. Нормативные аспекты клинического и исследовательского использования транскраниальной стимуляции постоянным током (tDCS): обзор и рекомендации группы экспертов. Clin Res Regul Aff. 2015 1 марта; 32(1):22-35.
13. Soterix Medical, Inc. получает одобрение CE Mark для терапии депрессии 1×1 tDCS | soterixmedical.com
14. Нисида К., Кошикава Ю., Морисима Ю. и др. Предстимульная активность мозга связана с изменением состояния тревожности при однократной транскраниальной стимуляции постоянным током. Передний шум нейронов. 2019;13:266. doi:10.3389/fnhum.2019.00266
15. Клэнси К.Дж. и др. Длительное увеличение связи и снижение беспокойства за счет транскраниальной стимуляции переменным током. соц. Познан. Оказывать воздействие. Неврологи.
Френьи Ф., Ницше М.А., Лоо К.К., Брунони А.Р., Маранголо П., Лейте Дж., Карвалью С., Болоньини Н., Каумо В., Пайк Н.Дж., Симис М., Уеда К., Эхтиари Х., Луу П., Такер Д.М., Тайлер В.Дж., Brunelin J, Datta A, Juan CH, Venkatasubramanian G, Boggio PM, Bikson M. Нормативные аспекты клинического и исследовательского использования транскраниальной стимуляции постоянным током (tDCS): обзор и рекомендации группы экспертов. Clin Res Regul Aff. 2015 1 марта; 32(1):22-35.
13. Soterix Medical, Inc. получает одобрение CE Mark для терапии депрессии 1×1 tDCS | soterixmedical.com
14. Нисида К., Кошикава Ю., Морисима Ю. и др. Предстимульная активность мозга связана с изменением состояния тревожности при однократной транскраниальной стимуляции постоянным током. Передний шум нейронов. 2019;13:266. doi:10.3389/fnhum.2019.00266
15. Клэнси К.Дж. и др. Длительное увеличение связи и снижение беспокойства за счет транскраниальной стимуляции переменным током. соц. Познан. Оказывать воздействие. Неврологи. 2018;13:1305–1316.
16. Неврологические устройства; Реклассификация краниальных электротерапевтических стимуляторов, предназначенных для лечения тревоги и/или бессонницы; Дата вступления в силу требования о предварительном одобрении краниальных электротерапевтических стимуляторов, предназначенных для лечения депрессии | Federalregister.gov
17. Коффман Б., Трамбо М., Флорес Р., Гарсия К., ван дер Мерве А., Вассерманн Э., Вайзенд М. и Кларк В., 2020. Влияние Tdcs на Производительность и обучение обнаружению целей: взаимодействие с характеристиками стимула и экспериментальным планом.
18. Андре Р. Брунони, доктор медицины, доктор философии, Адриано Х. Моффа, психолог, Бернардо Сампайо-Джуниор, доктор медицины, Лукас Боррионе, доктор медицины, Марина Л. Морено, психолог, Ракель А. Фернандес , Psy.D., Беатрис П. Веронези, Psy.D., Барбара С. Ногейра, Psy.D., Луана В.М. Апарисио, доктор медицины, Лаис Б. Разза, психолог, Ренан Чаморро, психолог, Луара С. Торт. Испытание электрической терапии постоянным током в сравнении с эсциталопрамом при депрессии
Список авторов.
2018;13:1305–1316.
16. Неврологические устройства; Реклассификация краниальных электротерапевтических стимуляторов, предназначенных для лечения тревоги и/или бессонницы; Дата вступления в силу требования о предварительном одобрении краниальных электротерапевтических стимуляторов, предназначенных для лечения депрессии | Federalregister.gov
17. Коффман Б., Трамбо М., Флорес Р., Гарсия К., ван дер Мерве А., Вассерманн Э., Вайзенд М. и Кларк В., 2020. Влияние Tdcs на Производительность и обучение обнаружению целей: взаимодействие с характеристиками стимула и экспериментальным планом.
18. Андре Р. Брунони, доктор медицины, доктор философии, Адриано Х. Моффа, психолог, Бернардо Сампайо-Джуниор, доктор медицины, Лукас Боррионе, доктор медицины, Марина Л. Морено, психолог, Ракель А. Фернандес , Psy.D., Беатрис П. Веронези, Psy.D., Барбара С. Ногейра, Psy.D., Луана В.М. Апарисио, доктор медицины, Лаис Б. Разза, психолог, Ренан Чаморро, психолог, Луара С. Торт. Испытание электрической терапии постоянным током в сравнении с эсциталопрамом при депрессии
Список авторов.