
Электротехника
Электротехника
ОглавлениеПРЕДИСЛОВИЕВВЕДЕНИЕ ГЛАВА I. ЛИНЕЙНЫЕ ЦЕПИ ПЕРЕМЕННОГО ТОКА 1. ОДНОФАЗНЫЕ ЦЕПИ § 1.2. ПРИНЦИП ПОЛУЧЕНИЯ ПЕРЕМЕННОЙ СИНУСОИДАЛЬНОЙ ЭДС § 1.3. ДЕЙСТВУЮЩИЕ ЗНАЧЕНИЯ ТОКА И НАПРЯЖЕНИЯ § 1.4. СРЕДНЕЕ ЗНАЧЕНИЕ ПЕРЕМЕННОГО ТОКА  5. МЕТОД ВЕКТОРНЫХ ДИАГРАММ 5. МЕТОД ВЕКТОРНЫХ ДИАГРАММ§ 1.6. СОПРОТИВЛЕНИЯ В ЦЕПЯХ ПЕРЕМЕННОГО ТОКА § 1.7. ЦЕПЬ ПЕРЕМЕННОГО ТОКА С АКТИВНЫМ СОПРОТИВЛЕНИЕМ § 1.8. ЦЕПЬ ПЕРЕМЕННОГО ТОКА С ИНДУКТИВНОСТЬЮ § 1.9. ЦЕПЬ ПЕРЕМЕННОГО ТОКА С АКТИВНО-ИНДУКТИВНОЙ НАГРУЗКОЙ § 1.10. ЦЕПЬ ПЕРЕМЕННОГО ТОКА С ЕМКОСТЬЮ § 1.11. ЦЕПЬ ПЕРЕМЕННОГО ТОКА С АКТИВНО-ЕМКОСТНОЙ НАГРУЗКОЙ § 1.12. ПОСЛЕДОВАТЕЛЬНОЕ СОЕДИНЕНИЕ R, L И С. КОЭФФИЦИЕНТ МОЩНОСТИ § 1.13. РЕЗОНАНС НАПРЯЖЕНИЙ § 1.14. РЕЗОНАНС ТОКОВ § 1.15. СПОСОБЫ ПОВЫШЕНИЯ КОЭФФИЦИЕНТА МОЩНОСТИ § 1.16. ПРОВОДИМОСТЬ И РАСЧЕТ ЭЛЕКТРИЧЕСКИХ ЦЕПЕЙ § 1.17. СИМВОЛИЧЕСКИЙ МЕТОД 2. ТРЕХФАЗНЫЕ ЦЕПИ § 1.19. ПРИНЦИП ПОСТРОЕНИЯ ТРЕХФАЗНОЙ СИСТЕМЫ § 1.21. СОЕДИНЕНИЕ ТРЕУГОЛЬНИКОМ § 1.22. МОЩНОСТЬ ТРЕХФАЗНОЙ СИСТЕМЫ ГЛАВА II. ЭЛЕКТРИЧЕСКИЕ ИЗМЕРЕНИЯ И ПРИБОРЫ § 2.3. ПОГРЕШНОСТИ ЭЛЕКТРИЧЕСКИХ ИЗМЕРЕНИЙ § 2.4. ОСНОВНЫЕ ДЕТАЛИ ЭЛЕКТРОИЗМЕРИТЕЛЬНЫХ ПРИБОРОВ § 2.5. МАГНИТОЭЛЕКТРИЧЕСКИЕ ПРИБОРЫ § 2.  6. ЭЛЕКТРОМАГНИТНЫЕ ПРИБОРЫ 6. ЭЛЕКТРОМАГНИТНЫЕ ПРИБОРЫ§ 2.7. ЭЛЕКТРОДИНАМИЧЕСКИЕ ПРИБОРЫ § 2.8. ФЕРРОДИНАМИЧЕСКИЕ ПРИБОРЫ § 2.9. ЭЛЕКТРОДИНАМИЧЕСКИЕ ВАТТМЕТРЫ § 2.10. ОДНОФАЗНЫЙ ФАЗОМЕТР § 2.11. ОДНОФАЗНЫЙ ИНДУКЦИОННЫЙ СЧЕТЧИК ЭЛЕКТРИЧЕСКОЙ ЭНЕРГИИ § 2.12. ОММЕТРЫ § 2.13. ЛОГОМЕТРЫ § 2.14. ТЕРМОЭЛЕКТРИЧЕСКИЕ ПРИБОРЫ § 2.15. ДЕТЕКТОРНЫЕ ПРИБОРЫ § 2.17. ПОНЯТИЕ О ЦИФРОВЫХ ИЗМЕРИТЕЛЬНЫХ ПРИБОРАХ § 2.18. ИЗМЕРЕНИЕ МОЩНОСТИ ТРЕХФАЗНОЙ СИСТЕМЫ § 2.19. ИЗМЕРЕНИЕ ЭНЕРГИИ ТРЕХФАЗНОЙ СИСТЕМЫ § 2.20. СПОСОБЫ ОПРЕДЕЛЕНИЯ КОЭФФИЦИЕНТА МОЩНОСТИ § 2.21. ПОНЯТИЕ ОБ ИЗМЕРЕНИЯХ НЕЭЛЕКТРИЧЕСКИХ ВЕЛИЧИН ЭЛЕКТРИЧЕСКИМИ МЕТОДАМИ ГЛАВА III. ТРАНСФОРМАТОРЫ § 3.2. УСТРОЙСТВО И ПРИНЦИП РАБОТЫ ТРАНСФОРМАТОРА § 3.3. ХОЛОСТОЙ РЕЖИМ РАБОТЫ ТРАНСФОРМАТОРА § 3.4. РАБОЧИЙ РЕЖИМ ТРАНСФОРМАТОРА § 3.5. КОЭФФИЦИЕНТ ПОЛЕЗНОГО ДЕЙСТВИЯ ТРАНСФОРМАТОРА § 3.6. ТРЕХФАЗНЫЕ ТРАНСФОРМАТОРЫ § 3.7. КОНСТРУКЦИИ ТРАНСФОРМАТОРОВ  8. АВТОТРАНСФОРМАТОР 8. АВТОТРАНСФОРМАТОР§ 3.9. ИЗМЕРИТЕЛЬНЫЕ ТРАНСФОРМАТОРЫ Глава IV. НЕЛИНЕЙНЫЕ ЭЛЕКТРИЧЕСКИЕ ЦЕПИ. ВЫПРЯМИТЕЛИ 4.1. ХАРАКТЕРИСТИКИ НЕЛИНЕЙНЫХ ЭЛЕКТРИЧЕСКИХ ЦЕПЕЙ И ЭЛЕМЕНТОВ § 4.2. ПОЛУПРОВОДНИКОВЫЕ ДИОДЫ § 4.3. ТИРИСТОРЫ § 4.4. ОСНОВНЫЕ СХЕМЫ ВЫПРЯМЛЕНИЯ ПЕРЕМЕННОГО ТОКА § 4.5. ПРИМЕНЕНИЕ ТИРИСТОРОВ ДЛЯ ВЫПРЯМЛЕНИЯ И РЕГУЛИРОВАНИЯ ТОКА § 4.6. СГЛАЖИВАЮЩИЕ ФИЛЬТРЫ § 4.7. ПОНЯТИЕ ОБ ИНВЕРТОРАХ § 4.8. ФЕРРОРЕЗОНАНС В НЕЛИНЕЙНЫХ ЭЛЕКТРИЧЕСКИХ ЦЕПЯХ § 4.9. ФЕРРОРЕЗОНАНСНЫЙ СТАБИЛИЗАТОР НАПРЯЖЕНИЯ § 4.10. ШКОЛЬНЫЕ ВЫПРЯМИТЕЛИ ГЛАВА V. МАШИНЫ ПЕРЕМЕННОГО ТОКА § 5.2. ПРИНЦИП РАБОТЫ И УСТРОЙСТВО АСИНХРОННОГО ДВИГАТЕЛЯ § 5.3. СОЗДАНИЕ ВРАЩАЮЩЕГОСЯ МАГНИТНОГО ПОЛЯ ТРЕХФАЗНОЙ СИСТЕМОЙ § 5.4. СКОРОСТЬ ВРАЩЕНИЯ МАГНИТНОГО ПОЛЯ. ТИПЫ ОБМОТОК СТАТОРА § 5.5. СКОЛЬЖЕНИЕ АСИНХРОННЫХ ДВИГАТЕЛЕЙ § 1.6. МАГНИТНЫЙ ПОТОК ЭДС И ТОКИ АСИНХРОННОГО ДВИГАТЕЛЯ § 5.7. ВЕКТОРНАЯ ДИАГРАММА АСИНХРОННОГО ДВИГАТЕЛЯ § 5.  8. АСИНХРОННЫЙ ДВИГАТЕЛЬ С КОНТАКТНЫМИ КОЛЬЦАМИ 8. АСИНХРОННЫЙ ДВИГАТЕЛЬ С КОНТАКТНЫМИ КОЛЬЦАМИ§ 5.9. РАБОЧИЕ ХАРАКТЕРИСТИКИ АСИНХРОННОГО ДВИГАТЕЛЯ § 5.10. ПУСК В ХОД АСИНХРОННЫХ ДВИГАТЕЛЕЙ § 5.11. РЕВЕРСИРОВАНИЕ И РЕГУЛИРОВАНИЕ СКОРОСТИ АСИНХРОННЫХ ДВИГАТЕЛЕЙ § 5.13. ПРИМЕНЕНИЕ ТРЕХФАЗНЫХ АСИНХРОННЫХ ДВИГАТЕЛЕЙ § 5.14. УСТРОЙСТВО И ПРИНЦИП РАБОТЫ СИНХРОННОГО ГЕНЕРАТОРА § 5.15. ЭДС СИНХРОННОГО ГЕНЕРАТОРА § 5.16. РЕАКЦИЯ ЯКОРЯ § 5.17. ОСНОВНЫЕ ХАРАКТЕРИСТИКИ СИНХРОННОГО ГЕНЕРАТОРА § 5.18. УПРОЩЕННАЯ ВЕКТОРНАЯ ДИАГРАММА СИНХРОННОГО ГЕНЕРАТОРА § 5.19. РАБОТА СИНХРОННОГО ГЕНЕРАТОРА ПАРАЛЛЕЛЬНО С СЕТЬЮ § 5.20. ОБРАТИМОСТЬ СИНХРОННЫХ МАШИН. ПРИНЦИП РАБОТЫ СИНХРОННОГО ДВИГАТЕЛЯ § 5.21. ПУСК И ОСТАНОВКА СИНХРОННОГО ДВИГАТЕЛЯ § 5.22. ВЛИЯНИЕ ТОКА ВОЗБУЖДЕНИЯ НА РАБОТУ СИНХРОННОГО ДВИГАТЕЛЯ. СИНХРОННЫЙ КОМПЕНСАТОР § 5.23. РЕАКТИВНЫЕ СИНХРОННЫЕ ДВИГАТЕЛИ § 5.24. ПРИМЕНЕНИЕ СИНХРОННЫХ ДВИГАТЕЛЕЙ § 6.  2. ПРИНЦИП РАБОТЫ И УСТРОЙСТВО ГЕНЕРАТОРА ПОСТОЯННОГО ТОКА. ТИПЫ ОБМОТОК ЯКОРЯ 2. ПРИНЦИП РАБОТЫ И УСТРОЙСТВО ГЕНЕРАТОРА ПОСТОЯННОГО ТОКА. ТИПЫ ОБМОТОК ЯКОРЯ§ 6.3. ЭДС И ЭЛЕКТРОМАГНИТНЫЙ МОМЕНТ ГЕНЕРАТОРА ПОСТОЯННОГО ТОКА § 6.4. РЕАКЦИЯ ЯКОРЯ § 6.5. КОММУТАЦИЯ § 6.6. СПОСОБЫ ВОЗБУЖДЕНИЯ ГЕНЕРАТОРОВ ПОСТОЯННОГО ТОКА § 6.7. ОБРАТИМОСТЬ МАШИН ПОСТОЯННОГО ТОКА. ДВИГАТЕЛИ § 6.8. ДВИГАТЕЛЬ ПАРАЛЛЕЛЬНОГО И НЕЗАВИСИМОГО ВОЗБУЖДЕНИЯ § 6.9. ДВИГАТЕЛЬ ПОСЛЕДОВАТЕЛЬНОГО ВОЗБУЖДЕНИЯ § 6.10. ДВИГАТЕЛЬ СМЕШАННОГО ВОЗБУЖДЕНИЯ § 6.11. КОЛЛЕКТОРНЫЕ ДВИГАТЕЛИ ПЕРЕМЕННОГО ТОКА ГЛАВА VII. ЭЛЕМЕНТЫ АВТОМАТИКИ § 7.3. ДАТЧИКИ § 7.4. АВТОМАТИЧЕСКИЙ КОНТРОЛЬ § 7.5. АВТОМАТИЧЕСКОЕ УПРАВЛЕНИЕ § 7.6. АВТОМАТИЧЕСКОЕ РЕГУЛИРОВАНИЕ § 7.7. ТЕЛЕМЕХАНИКА § 7.8. КОМПЛЕКСНАЯ АВТОМАТИЗАЦИЯ ГЛАВА VIII. ПРОИЗВОДСТВО, ПЕРЕДАЧА И ИСПОЛЬЗОВАНИЕ ЭЛЕКТРИЧЕСКОЙ ЭНЕРГИИ В НАРОДНОМ ХОЗЯЙСТВЕ § 8.1. ЭЛЕКТРИЧЕСКИЕ СТАНЦИИ § 8.2. ЭНЕРГЕТИЧЕСКИЕ СИСТЕМЫ § 8.3. ПЕРЕДАЧА ЭЛЕКТРИЧЕСКОЙ ЭНЕРГИИ ПОСТОЯННЫМ ТОКОМ § 8.4. АВТОМАТИЗАЦИЯ ЭЛЕКТРИЧЕСКИХ СТАНЦИЙ И ПОДСТАНЦИЙ ГЛАВА IX.  СОВРЕМЕННЫЕ ТЕНДЕНЦИИ РАЗВИТИЯ ЭЛЕКТРОЭНЕРГЕТИКИ СОВРЕМЕННЫЕ ТЕНДЕНЦИИ РАЗВИТИЯ ЭЛЕКТРОЭНЕРГЕТИКИГЛАВА X. ЭЛЕМЕНТЫ ТЕХНИКИ БЕЗОПАСНОСТИ § 10.2. ЭЛЕКТРИЧЕСКОЕ СОПРОТИВЛЕНИЕ ТЕЛА ЧЕЛОВЕКА § 10.3. ОСНОВНЫЕ ПРИЧИНЫ ПОРАЖЕНИЯ ЭЛЕКТРИЧЕСКИМ ТОКОМ § 10.4. ЗАЩИТНОЕ ЗАЗЕМЛЕНИЕ И ЗАЗЕМЛЕНИЕ НА НЕЙТРАЛЬ (ЗАНУЛЕНИЕ) § 10.5. ЗАЩИТНЫЕ СРЕДСТВА И КОНТРОЛЬ СОСТОЯНИЯ ИЗОЛЯЦИИ ЭЛЕКТРОУСТАНОВОК § 10.6. ОКАЗАНИЕ ПЕРВОЙ ПОМОЩИ ПОРАЖЕННОМУ ЭЛЕКТРИЧЕСКИМ ТОКОМ § 10.7. ОСНОВНЫЕ ПРАВИЛА ТЕХНИКИ БЕЗОПАСНОСТИ В УЧЕБНЫХ ЛАБОРАТОРИЯХ ЛИТЕРАТУРА |
Метод индукционного каротажного зондирования (ИКЗ)
Метод индукционного каротажного зондирования (ИКЗ)
Истории успешного применения технологий
Индукционным каротажным зондированием (ИКЗ) называют изучение удельного сопротивления (удельной электропроводности) пересеченных скважиной горных пород, основанное на измерении вторичного поля вихревых токов, индуцированных в породе. В основе метода лежит закон электромагнитной индукции (закон Фарадея), устанавливающий взаимосвязь между магнитными и электрическими явлениями. Вертикальная разрешающая способность метода – 35 см, горизонтальная разрешающая способность метода – 250 см.
В основе метода лежит закон электромагнитной индукции (закон Фарадея), устанавливающий взаимосвязь между магнитными и электрическими явлениями. Вертикальная разрешающая способность метода – 35 см, горизонтальная разрешающая способность метода – 250 см.
Типовые условия применения метода:
- применяется в необсаженных скважинах, заполненных промывочной жидкостью любой минерализации или на нефтяной основе без содержания магнитных добавок, а также сухих скважинах;
- может применяться в скважинах, обсаженных токонепроводящими колоннами;
- ограничением в применении метода являются высокое содержание в промывочной жидкости компонентов с сильными магнитными свойствами и значения удельного электрического сопротивления пород, превышающие 500 Ом*м.
| ПРИМЕНЕНИЕ: |
|---|
|

ТЕХНИЧЕСКИЕ ОСОБЕННОСТИ:
| Прибор 4ИК-Т-76 |
|
| Прибор 4ИК-1Т |
|
| Прибор 5ИК-Т-76 |
|

|
Обозначение прибора:
|
Регистрируемые параметры:
|
Единицы измерения:
| |
ТЕХНИЧЕСКИЕ ХАРАКТЕРИСТИКИ
|
|
Прибор 4ИК-Т-76 |
Прибор 4ИК-1Т |
Прибор 5ИК-Т-76 |
|---|---|---|---|
|
Длина, м |
3,95 |
5,35 |
3,95 |
|
Диаметр, мм |
76 |
76 |
76 |
|
Масса, кг |
40 |
80 |
40 |
|
Максимальная рабочая температура, °С |
120 |
175 |
120 |
|
Максимальное рабочее давление, МПа |
80 |
140 |
80 |
|
Максимальная скорость записи, м/ч в интервале М 1:200 (детальных исследований) |
1800 |
2000 |
1800 |
|
Диаметр исследуемых скважин, мм |
от 110
|
от 90
|
от 110
|
|
Положение в скважине |
свободное/
|
свободное/
|
свободное/
|
|
Комбинируемость |
транзитный |
транзитный |
транзитный |
|
Радиус исследования |
|
|
|
|
зонд 3И2. |
2,97 |
2,97 |
2,97 |
|
зонд 3И1.26 |
1,82 |
1,82 |
1,82 |
|
зонд 3И0.85 |
1,23 |
1,23 |
1,23 |
|
зонд 3И0.5 |
0,72 |
0,72 |
0,72 |
|
зонд 3И0. |
— |
— |
0,4 |
 05
05 3
3ПРИНЦИПИАЛЬНАЯ КОНСТРУКЦИЯ МОДУЛЯ
Прибор 5ИК-Т-76 содержит 5 (4, в случае с прибором 4ИК-Т-76) трехкатушечных индукционных зондов, каждый из которых имеет 2 генераторные (основную и компенсационную) и одну (общую для всех зондов) измерительную катушку L1. Генераторные катушки совместно с измерительной образуют следующие зонды:
L2, L3, L1 – зонд 3И0.3, питается генератором U1;
L4, L5, L1 – зонд 3И0.5, питается генератором U2;
L6, L7, L1 – зонд 3И0.85, питается генератором U3;
L8, L9, L1 – зонд 3И1.26, питается генератором U4;
L10, L11, L1 – зонд 3И2.05, питается генератором U5.
Назад в раздел
Пинчук Михаил Михайлович
Главный инженер
телефон: 8 (34667)4-41-84
mpinchuk@kngf. org
org
Борискин Виктор Алексеевич Начальник КИП-1 телефон: 8 (34667) 4-47-19 [email protected]
Рыбин Никита Петрович Заместитель генерального директора по производству телефон: 8 (34667) 4-44-78 [email protected]
Кокоулин Сергей Викторович Главный инженер Ямальской промыслово-геофизической экспедиции телефон: 8-908-89-55-729 [email protected]
Вопросы и ответы
Вопросов пока нет
Задать вопрос
ФИО* Компания Должность E-mail* Текст сообщения** — обязательные поля
Передача сигналов на большие расстояния с помощью потенциалов действия — нейробиология
NCBI Книжная полка. Служба Национальной медицинской библиотеки, Национальных институтов здоровья.
Purves D, Augustine GJ, Fitzpatrick D и др., редакторы. Неврология. 2-е издание. Сандерленд (Массачусетс): Sinauer Associates; 2001.
- По соглашению с издателем эта книга доступна через функцию поиска, но не может быть просмотрена.
Показать детали
Критерий поиска
Зависящие от напряжения механизмы генерации потенциала действия также объясняют передачу этих электрических сигналов на большие расстояния. Вспомним из главы 2, что нейроны являются относительно плохими проводниками электричества, по крайней мере, по сравнению с проводом. Проведение тока по проводам и по нейронам при отсутствии потенциалов действия называется пассивным течением тока (вставка C). Пассивные электрические свойства аксона нервной клетки можно определить, измерив изменение напряжения в результате импульса тока, прошедшего через мембрану аксона. Если этот импульс тока недостаточно велик, чтобы генерировать потенциалы действия, величина результирующего изменения потенциала экспоненциально затухает с увеличением расстояния от места введения тока (). Как правило, потенциал падает до небольшой доли своего начального значения на расстоянии не более пары миллиметров от места инъекции (). Прогрессирующее уменьшение амплитуды индуцированного изменения потенциала происходит из-за просачивания введенного тока через аксональную мембрану; соответственно, меньше тока доступно для изменения мембранного потенциала дальше по аксону. Таким образом, негерметичность аксональной мембраны препятствует эффективной пассивной передаче электрических сигналов во всех, кроме самых коротких аксонов (длиной 1 мм и менее). Точно так же негерметичность мембраны замедляет ход ответов, измеренных на увеличивающихся расстояниях от места, где вводился ток (10).
Как правило, потенциал падает до небольшой доли своего начального значения на расстоянии не более пары миллиметров от места инъекции (). Прогрессирующее уменьшение амплитуды индуцированного изменения потенциала происходит из-за просачивания введенного тока через аксональную мембрану; соответственно, меньше тока доступно для изменения мембранного потенциала дальше по аксону. Таким образом, негерметичность аксональной мембраны препятствует эффективной пассивной передаче электрических сигналов во всех, кроме самых коротких аксонов (длиной 1 мм и менее). Точно так же негерметичность мембраны замедляет ход ответов, измеренных на увеличивающихся расстояниях от места, где вводился ток (10).
Box C
Свойства пассивной мембраны.
Рисунок 3.10
Пассивный ток в аксоне. (A) Экспериментальная установка для изучения локального потока электрического тока в аксоне. Электрод с током создает подпороговое изменение мембранного потенциала, который пассивно распространяется по аксону. (B) (подробнее…)
(B) (подробнее…)
Если повторить эксперимент, показанный на рис., с достаточно большим импульсом деполяризующего тока, чтобы вызвать потенциал действия, результат будет кардинально другим (). При этом потенциал действия возникает без декремента по всей длине аксона, что может быть на расстоянии метра и более (). Таким образом, потенциалы действия каким-то образом обходят присущую нейронам негерметичность.
Рисунок 3.11
Распространение потенциала действия. (A) Экспериментальная схема: электрод вызывает потенциал действия, вводя сверхпороговый ток. (B) потенциальные ответы, записанные в местах, указанных микроэлектродами. Амплитуда действия (подробнее…)
Как же тогда потенциалы действия проходят большие расстояния по такому плохому пассивному проводнику? Ответ отчасти дает наблюдение, что амплитуда потенциалов действия, регистрируемых на разных расстояниях, постоянна. Это поведение типа «все или ничего» указывает на то, что в распространении потенциала действия должно участвовать нечто большее, чем просто пассивный поток тока. Вторая подсказка возникает при изучении времени возникновения потенциалов действия, зарегистрированных на разных расстояниях от места стимуляции: потенциалы действия появляются позже и позже на больших расстояниях вдоль аксона (1). Таким образом, потенциал действия имеет измеримую скорость передачи, называемую скоростью проводимости. Задержка прихода потенциала действия в последовательно более удаленные точки вдоль аксона отличается от случая, показанного на рис. 2, в котором электрические изменения, вызванные протеканием пассивного тока, происходят более или менее в одно и то же время в последовательных точках.
Вторая подсказка возникает при изучении времени возникновения потенциалов действия, зарегистрированных на разных расстояниях от места стимуляции: потенциалы действия появляются позже и позже на больших расстояниях вдоль аксона (1). Таким образом, потенциал действия имеет измеримую скорость передачи, называемую скоростью проводимости. Задержка прихода потенциала действия в последовательно более удаленные точки вдоль аксона отличается от случая, показанного на рис. 2, в котором электрические изменения, вызванные протеканием пассивного тока, происходят более или менее в одно и то же время в последовательных точках.
Механизм распространения потенциала действия легко понять, если понять, как генерируются потенциалы действия и как ток пассивно течет по аксону (). Деполяризующий стимул — синаптический потенциал или рецепторный потенциал в интактном нейроне или введенный импульс тока в эксперименте — локально деполяризует аксон, тем самым открывая чувствительные к напряжению каналы Na + в этой области. Открытие каналов Na + вызывает движение Na + 9 внутрь.0034, и результирующая деполяризация мембранного потенциала генерирует потенциал действия в этом месте. Часть локального тока, генерируемого потенциалом действия, будет затем пассивно течь вниз по аксону таким же образом, как подпороговые токи распространяются по аксону (см. Обратите внимание, что этот пассивный ток не требует движения Na + вдоль аксона, а вместо этого происходит за счет перемещения заряда, подобно тому, что происходит, когда провода пассивно проводят электричество. Этот пассивный поток тока деполяризует мембранный потенциал в соседней области аксона, тем самым открывая Na + канала в соседней мембране. Локальная деполяризация запускает потенциал действия в этой области, который затем снова распространяется в непрерывном цикле, пока не будет достигнут конец аксона. Таким образом, распространение потенциала действия требует скоординированного действия двух форм протекания тока — пассивного протекания тока, а также активных токов, протекающих через потенциалзависимые ионные каналы.
Открытие каналов Na + вызывает движение Na + 9 внутрь.0034, и результирующая деполяризация мембранного потенциала генерирует потенциал действия в этом месте. Часть локального тока, генерируемого потенциалом действия, будет затем пассивно течь вниз по аксону таким же образом, как подпороговые токи распространяются по аксону (см. Обратите внимание, что этот пассивный ток не требует движения Na + вдоль аксона, а вместо этого происходит за счет перемещения заряда, подобно тому, что происходит, когда провода пассивно проводят электричество. Этот пассивный поток тока деполяризует мембранный потенциал в соседней области аксона, тем самым открывая Na + канала в соседней мембране. Локальная деполяризация запускает потенциал действия в этой области, который затем снова распространяется в непрерывном цикле, пока не будет достигнут конец аксона. Таким образом, распространение потенциала действия требует скоординированного действия двух форм протекания тока — пассивного протекания тока, а также активных токов, протекающих через потенциалзависимые ионные каналы. Регенеративные свойства открытия канала Na + позволяют потенциалам действия распространяться по принципу «все или ничего», действуя как усилитель в каждой точке вдоль аксона, тем самым обеспечивая передачу электрических сигналов на большие расстояния.
Регенеративные свойства открытия канала Na + позволяют потенциалам действия распространяться по принципу «все или ничего», действуя как усилитель в каждой точке вдоль аксона, тем самым обеспечивая передачу электрических сигналов на большие расстояния.
Рисунок 3.12
Для проведения потенциала действия требуется как активный, так и пассивный ток. Деполяризация в одной точке вдоль аксона локально открывает каналы Na + (точка 1) и вызывает потенциал действия в этой точке (А) аксона (момент времени t = 1). В результате внутрь (далее…)
По согласованию с издателем эта книга доступна через поиск, но не может быть просмотрена.
Copyright © 2001, Sinauer Associates, Inc.
Идентификатор книжной полки: NBK11107
- CITE на этой стр.
- Отключить глянцевые ссылки
Связанная информация
- PubMed
Ссылка на PubMed
.
 пусто.
пусто.Запись активности отключена.
Включить запись
Подробнее…
Потенциал действия
Главы Lodish 4-е издание: Глава 21 стр. 921 — 924
Мойес и Шульте: Глава 5 стр. 146-164
Активная проводимость нерв
— не очень эффективен, если нужно быстро передать сигнал в течение длительного времени расстояние
— сигнал уменьшается на расстоянии
— активная проводимость (т.е. генерация потенциала действия) означает, что сигнал проходит вдоль нерва без потери амплитуды
Рисунок 21-2. Lodish 4-е издание (а) Потенциал действия — это внезапный,
преходящая деполяризация мембраны с последующей реполяризацией
потенциал покоя около -60 мВ. Эта запись аксональной мембраны
потенциал в пресинаптическом нейроне показывает, что он производит одно действие
потенциала примерно каждые 4 миллисекунды. (б) Мембранный потенциал на
плазматическая мембрана пресинаптического нейрона измеряется небольшим электродом
вставлен в него. Потенциалы действия распространяются по аксону со скоростью до 100
метров в секунду. Их прибытие в синапс вызывает высвобождение
нейротрансмиттеры, которые связываются с рецепторами в постсинаптических клетках, обычно
деполяризуя мембрану (делая потенциал менее отрицательным) и стремясь
индуцировать в нем потенциал действия.
(б) Мембранный потенциал на
плазматическая мембрана пресинаптического нейрона измеряется небольшим электродом
вставлен в него. Потенциалы действия распространяются по аксону со скоростью до 100
метров в секунду. Их прибытие в синапс вызывает высвобождение
нейротрансмиттеры, которые связываются с рецепторами в постсинаптических клетках, обычно
деполяризуя мембрану (делая потенциал менее отрицательным) и стремясь
индуцировать в нем потенциал действия.
Потенциалы действия в нейронах в основном основаны на потенциалзависимых Na+ канал, некоторые нейроны используют как потенциалзависимый Na+-канал, так и потенциалзависимый K+ канал, некоторые нейроны используют только потенциалзависимый Na+ канал а некоторые нейроны используют потенциалзависимый Ca+2-канал
Мы будем использовать классический пример потенциала действия от гигантского аксона
кальмаров (беспозвоночных), а также потенциал действия, обнаруженный у немиелинизированных
аксонов млекопитающих. Этот потенциал действия состоит из двух компонентов: потенциалзависимого Na+
каналы и потенциалзависимые К+ каналы
Этот потенциал действия состоит из двух компонентов: потенциалзависимого Na+
каналы и потенциалзависимые К+ каналы
Канал Na+ с управляемым напряжением:
Канал имеет три состояния: закрытый, открытый и неактивный.
Closed to Open: Деполяризация необходима для открытия канала и
поэтому он действует, чтобы активировать себя в регенеративном цикле. Больше притока Na+
деполяризует мембрану, что открывает больше каналов, которые деполяризуют
мембрана больше.
Open to Inactive: Деполяризация также необходима для неактивного
канал. Как только канал будет открыт, он также переключится на неактивный
состоянии и не может быть открыт снова
От неактивного до закрытого: Канал не переключится обратно на закрытый
состоянии до тех пор, пока мембрана не реполяризуется (т. е. не вернется к исходному
мембранный потенциал покоя. Находясь в закрытом состоянии, его можно снова открыть
Канал K+ с управляемым напряжением (называемый каналом K+ с задержкой выпрямления)
Этот канал имеет только два состояния: закрытый и открытый.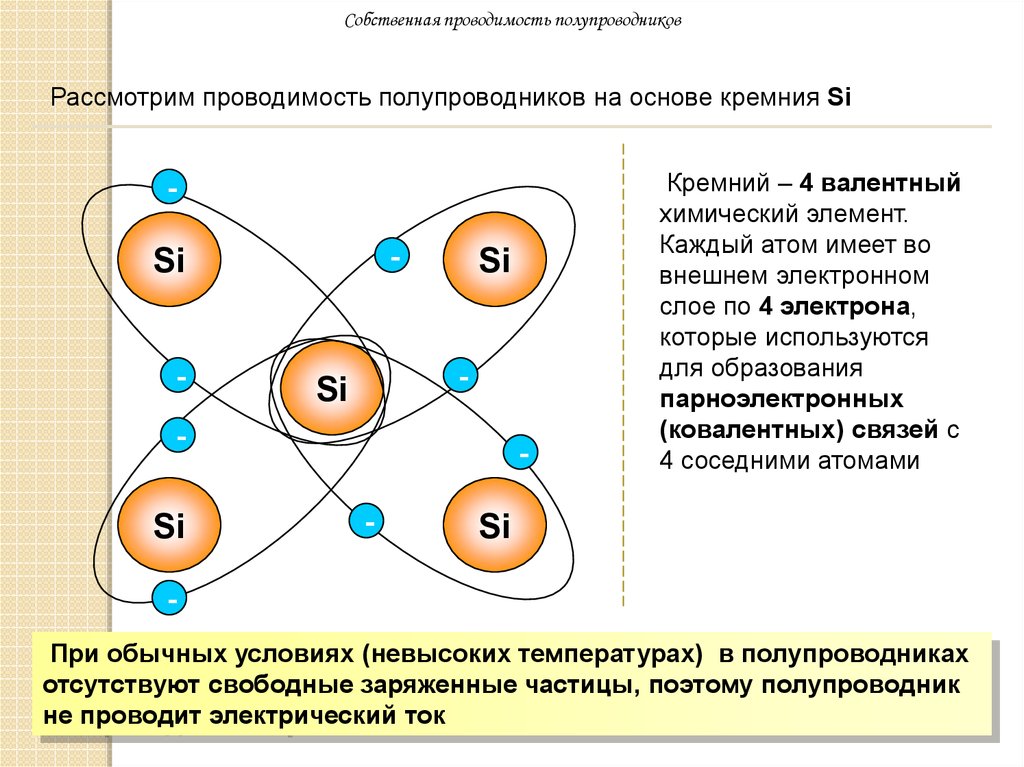
Closed to open: Канал открыт при сильной деполяризации,
тип, который вы обычно получаете в потенциале действия. Этот канал работает на
вернуть мембрану к потенциалу Нернста для K +, т.е. гиперполяризовать
мембрана
От открытия до закрытия: Канал закроется, когда мембрана станет
гиперполяризованные или реполяризованные. Поэтому этот канал работает, чтобы закрыть себя
вниз.
Компоненты потенциал действия
Рисунок 7-32 Lodish 5-е издание. Деполяризация плазматической мембраны вследствие открытие закрытых Na+ каналов. (а) Покоящиеся нейроны без гейтированных К+-каналов открыты, но более многочисленные закрытые Na+-каналы закрыты. Движение Ионы K+ наружу устанавливают внутри-отрицательный мембранный потенциал характерны для большинства клеток. (б) Открытие закрытых каналов Na+ позволяет приток достаточного количества ионов Na+ вызывает изменение мембранного потенциала.
Порог
— уровень деполяризации, необходимый для запуска потенциала действия (наиболее
нейроны имеют порог на уровне -50 мВ (т. е. деполяризация 10-15 мВ)
е. деполяризация 10-15 мВ)
— потенциал действия представляет собой событие «все или ничего», если нерв
амплитуда одного потенциала действия будет одинакова на всем протяжении нерва
не зависит от силы раздражителя.
— пороговое значение, отражающее необходимость срабатывания открытия потенциально управляемого
натриевый канал (для открытия требуется деполяризация примерно на 10–15 мВ)
Фаза повышения потенциала действия
— когда открываются натриевые каналы, ионы Na+ поступают в клетку, деполяризуя клетку открывается все больше и больше натриевых каналов = регенеративный ответ — регенеративный открытие натриевых каналов увеличивает мембранный потенциал до пика Равновесный потенциал Нернста для Na+
Пиковый потенциал действия
— во время потенциала действия мембранный потенциал приближается к потенциалу Нернста
равновесный потенциал для Na+
— по уравнению Гольдмана-Каца теперь доминирует проницаемость по Na+ (K+
и Cl- минорные компоненты) поэтому мембранный потенциал приближается к ENa
— обычно не дотягивает до ENa, меньше движущая сила на Na+ и начинаются каналы
быстро инактивироваться после активации
Падение фазы потенциала действия
— после достижения пика теперь потенциал действия падает, мембранный потенциал падает
назад к отдыху
— почему? Почему потенциал действия не остается вокруг ENa?
— две причины:
i) Na+-каналы переходят в неактивное состояние
ii) отсроченные K+-каналы открываются (гигантский аксон кальмара или немиелинизированные аксоны кальмара
позвоночные)
1) Инактивация Na+-каналов —
— Na+-каналы переходят в инактивированное состояние через 1-2 мс после первого открытия
— инактивирован = НЕ может быть повторно открыт
— поэтому мембранный потенциал теперь определяется в основном K+ (так же, как потенциал покоя) и мембрана начинает реполяризоваться
2) Каналы K+ с задержкой открыты (так называемый выпрямитель с задержкой; Na+ канал)
— открывается примерно через 1-2 мс пороговой деполяризации
— теперь K+ вытекает из клетки и ускоряет процесс реполяризации
— вызывает гиперполяризацию после потенциала действия, потому что открывается K+ каналы делают проницаемость K+ выше, чем в состоянии покоя, а мембраны более минус внутри-гиперполяризация мембраны вызывает закрытие K+ каналов
— затем мембрана возвращается в исходное положение
Реполяризация
— потенциалзависимые каналы Na+ и потенциалзависимые каналы K+ теперь закрыты, поэтому
мембрана возвращается в состояние покоя
— т. е. каналы утечки — единственные открытые каналы и снова устанавливают мембрану
потенциал
е. каналы утечки — единственные открытые каналы и снова устанавливают мембрану
потенциал
Рефрактерный период
— разделенный на две части
i) абсолютный рефрактерный период
ii) относительный рефрактерный период
1) Абсолютный рефрактерный период
— Na+ каналы неактивны и НЕ МОГУТ быть открыты, сколько бы
мембрана в это время деполяризована
— другой потенциал действия не может генерироваться в этой части нерва при
на этот раз
2) Относительный рефрактерный период
— когда мембрана реполяризуется = переходит к более отрицательным потенциалам, это вызывает
Na+-каналы переходят из неактивного состояния в закрытое.
— этому процессу способствует гиперполяризация путем открытия К+ каналов
— когда канал Na+ находится в закрытом состоянии, его можно снова открыть с помощью
деполяризация
— в течение относительного рефрактерного периода все больше и больше каналов Na+ доступны для
быть открытым и, следовательно, увеличить шансы запуска потенциала действия
Частота потенциалов действия
— Если потенциал действия, если все или ничего, как нерв передает
сила раздражителя?
— напр. как чувствительный нерв различает легкое прикосновение (перо)
и грубый абразивный штрих (наждачная бумага)?
как чувствительный нерв различает легкое прикосновение (перо)
и грубый абразивный штрих (наждачная бумага)?
— информация указана по частоте потенциалов действия вдоль
нерв.
— сила раздражителя (ввод тока в нерв экспериментально
введение большого тока или в реальной жизни по реакции сенсорного рецептора)
вызывает разную частоту потенциалов действия
— следовательно: легкое прикосновение — нечастые потенциалы действия; грубое прикосновение — больше
частые потенциалы действия
— рефрактерный период ограничивает частоту потенциала действия
— в течение относительного рефрактерного периода может генерироваться потенциал действия
но с повышенным порогом и уменьшенной амплитудой
— повышенный порог, потому что приходится преодолевать гиперполяризацию
— уменьшенная амплитуда, потому что для открытия доступно меньше каналов Na+ (многие
все еще в неактивном состоянии) и, таким образом, получить меньше Na+, поступающего в ячейку
(другими словами, проницаемость или проводимость Na+ уменьшается во время
относительный рефрактерный период — увеличивается к концу периода)
Потенциалы направления действия
Рисунок 21-14, Лодское 4-е издание ИЛИ Рисунок 7-35, Лодское 5-е издание. Однонаправленное проведение потенциала действия из-за переходного
инактивация потенциалзависимых Na+-каналов. В момент времени 0 потенциал действия
(фиолетовый) находится на расстоянии 2 мм от аксона. Деполяризация мембраны
пассивно распространяется в обоих направлениях вдоль аксона (рис. 21.11). Потому что
каналы Na+ в положении 1 мм все еще инактивированы (зеленые), они
еще не может быть вновь открыт из-за небольшой деполяризации, вызванной пассивным распространением.
Каждая область мембраны рефрактерна (неактивна) в течение нескольких миллисекунд.
после прохождения потенциала действия. Таким образом, деполяризация на 2-мм
сайт в момент времени 0 запускает потенциалы действия только вниз по течению; в 1 мс действие
потенциал проходит положение 3 мм, а через 2 мс возникает потенциал действия.
минуя положение 4 мм.
Однонаправленное проведение потенциала действия из-за переходного
инактивация потенциалзависимых Na+-каналов. В момент времени 0 потенциал действия
(фиолетовый) находится на расстоянии 2 мм от аксона. Деполяризация мембраны
пассивно распространяется в обоих направлениях вдоль аксона (рис. 21.11). Потому что
каналы Na+ в положении 1 мм все еще инактивированы (зеленые), они
еще не может быть вновь открыт из-за небольшой деполяризации, вызванной пассивным распространением.
Каждая область мембраны рефрактерна (неактивна) в течение нескольких миллисекунд.
после прохождения потенциала действия. Таким образом, деполяризация на 2-мм
сайт в момент времени 0 запускает потенциалы действия только вниз по течению; в 1 мс действие
потенциал проходит положение 3 мм, а через 2 мс возникает потенциал действия.
минуя положение 4 мм.
— рефрактерный период также определяет направление потенциала действия
— деполяризующий ток от потенциала действия может пассивно распространяться в
в любом направлении
— в одну сторону каналы Na+ находятся в закрытом состоянии и готовы к открытию,
поэтому распространяющийся ток может вызвать потенциал действия в этом
соседний регион
— в противном случае каналы Na+ находятся в неактивном состоянии и не могут быть
открыты, поэтому ток распространения не влияет на каналы в этом
региона и потенциал действия не является триггерным
Каналы и рецепторы
На лекциях мы познакомим вас с широким спектром ионных
каналы. Они варьируются от каналов утечки (K+, Na+, Cl- и т. д.),
ионные каналы (K+, Na+ и Ca+2 и т. д.) и выровненные закрытые ионные каналы (K+/Na+,
Cl- и др.).
Они варьируются от каналов утечки (K+, Na+, Cl- и т. д.),
ионные каналы (K+, Na+ и Ca+2 и т. д.) и выровненные закрытые ионные каналы (K+/Na+,
Cl- и др.).
Рисунок 21-8, Lodish, 4-е издание. Ионные каналы в плазматических мембранах нейронов.
Каждый тип канального белка выполняет определенную функцию в электрическом
активности нейронов. а) покоящиеся К+-каналы ответственны за генерацию
потенциал покоя через мембрану. (b) Потенциал-зависимые каналы
отвечает за распространение потенциалов действия вдоль аксональной мембраны. (с,
г) Два типа ионных каналов в дендритах и телах клеток отвечают за
для генерации электрических сигналов в постсинаптических клетках. Один тип (с) имеет
сайт для связывания специфического внеклеточного нейротрансмиттера (синий кружок).
Другой тип (d) связан с рецептором нейротрансмиттера через G
белок; он отвечает на внутриклеточные сигналы (красный кружок), индуцированные
связывание нейротрансмиттера с отдельным белком-рецептором (не показано). Сигналы, активирующие различные каналы, включают Ca2+, циклический GMP и Ga.
субъединицы тримерных G-белков (глава 20).
Сигналы, активирующие различные каналы, включают Ca2+, циклический GMP и Ga.
субъединицы тримерных G-белков (глава 20).
Большинство белков, из которых состоят различные типы ионных каналов, очень похожи по своей структуре и имеют консервативную аминокислотную последовательность. Этот степень сохранения происходит между различными типами каналов и через разновидность. Так, например, потенциалзависимый Na+-канал дрозофилы очень похож на потенциалозависимый Na+-канал человека и т. д. Все ионные каналы состоит из альфа-спиралей, которые охватывают липидный бислой. Те, что связываются с липидный бислой состоит из гидрофобных аминокислот (Phe, Ile, Leu и др.) которые охватывают около 20 аминокислот. Те альфа-спирали, которые выстилают поры, состоит из гидрофильных остатков, обеспечивающих поток ионов (Lys, Arg и т. д.).
Рисунок 21-27, Lodish 4-е издание. Предлагаемые трансмембранные структуры из четырех
Типы белков с закрытыми ионными каналами.
(а) Потенциал-зависимый К+-канал шейкера, выделенный из дрозофилы, представляет собой
тетрамер из четырех идентичных субъединиц, каждая из которых содержит 656 аминокислот и шесть
трансмембранные альфа-спирали (обозначены арабскими цифрами). Helix 4 (бордовый)
действует как датчик напряжения, а неспиральные сегменты P между спиралями 5
и 6 выстилают ионную пору. (b) Циклические AMP- или циклические GMP-управляемые ионные каналы
также содержат четыре субъединицы. Поскольку ни одна из трансмембранных альфа-спиралей
действуют как датчик напряжения, эти каналы не являются потенциалозависимыми. Скорее,
связывание цАМФ или цГМФ с цитозольным сегментом вызывает открытие этих
каналы, которых много в чувствительных клетках зрительного и
обонятельные системы (см. рис. 21-47). (c) Потенциалзависимый Na+ (и Ca+2)
каналы представляют собой мономерные белки, содержащие 1800–2000 аминокислот, организованные
на четыре гомологичных трансмембранных домена (обозначены римскими цифрами). Около 64 процентов остатков сходны или идентичны по последовательности в
оба канала. Считается, что каждый из четырех гомологичных доменов содержит шесть
трансмембранные альфа-спирали, сходные по структуре с таковыми в шейкере
канальные мономеры в (а).
Около 64 процентов остатков сходны или идентичны по последовательности в
оба канала. Считается, что каждый из четырех гомологичных доменов содержит шесть
трансмембранные альфа-спирали, сходные по структуре с таковыми в шейкере
канальные мономеры в (а).
Пора канала
Все рассматриваемые ионные каналы имеют общую черту. Пора, которая позволяет рассматриваемый ион (ионы) течет через липидный бислой. Пора специфична к определенному иону или ионам. Например, канал утечки K+ пропускает только ионы K+. протекать через мембрану.
Рисунок 7-16, Lodish, 5-е издание. Механизм ионной селективности и транспорта в покоящихся К+ каналах. (а) Схематическая диаграмма ионов K+ и Na+, гидратированных в растворе и в поре К+ канала. (б) Высокое разрешение карта электронной плотности, полученная с помощью рентгеновской кристаллографии, показывающая ионы K + прохождение через фильтр селективности.
Датчик напряжения
Датчик напряжения представляет собой альфа-спираль, расположенную в канале и охватывающую
мембрана. Датчик напряжения имеет положительный заряд на каждой третьей аминокислоте.
Датчик перемещается в ответ на деполяризацию (т.е. увеличение положительного
заряд на внутренней мембране вызывает физическое движение
датчик напряжения). Сенсорная альфа-спираль спрятана внутри белка канала.
(т.е. защищен от гидрофобного липидного двойного слоя остальной частью ионного канала
белок).
Датчик напряжения имеет положительный заряд на каждой третьей аминокислоте.
Датчик перемещается в ответ на деполяризацию (т.е. увеличение положительного
заряд на внутренней мембране вызывает физическое движение
датчик напряжения). Сенсорная альфа-спираль спрятана внутри белка канала.
(т.е. защищен от гидрофобного липидного двойного слоя остальной частью ионного канала
белок).
Рисунок 21-13 Lodish, 4-е издание ИЛИ Рисунок 7-33 Lodish, 5-е издание. Состав
и функция потенциалзависимого Na+ канала.
Как и все потенциалзависимые каналы, он содержит четыре трансмембранных домена,
каждый из которых вносит свой вклад в центральную пору, через которую движутся ионы.
критические компоненты, контролирующие движение ионов Na+, показаны на
вид в разрезе. а) в закрытом состоянии покоя ворота препятствуют
канал, ингибирующий движение Na+, а сегмент, инактивирующий канал,
свободно в цитозоле. Канальный белок содержит четыре чувствительных к напряжению альфа-канала. спирали (бордовые), каждая третья из которых имеет положительно заряженные боковые цепи
остаток. Притяжение этих зарядов к отрицательным внутренностям
покоящиеся клетки помогают держать канал закрытым. б) когда мембрана становится
деполяризованные (снаружи отрицательные), чувствительные к напряжению спирали движутся к
внешней поверхности плазматической мембраны, вызывая немедленное конформационное изменение
сегмент ворот, открывающий канал для поступления ионов Na+. (c) В течение
миллисекунды после открытия спирали, чувствительные к напряжению, возвращаются в исходное положение.
положение, и сегмент инактивации канала (фиолетовый) перемещается в открытый
канала, препятствуя дальнейшему движению ионов. Когда мембранный потенциал
перевернутый так, что внутренняя часть снова отрицательная, ворота перемещаются обратно в
положение блокировки (не показано). Через 1 2 мс канал деактивируется.
сегмент вытесняется из отверстия канала, и белок возвращается к
закрытое состояние покоя (а), где он может быть снова открыт путем деполяризации.
спирали (бордовые), каждая третья из которых имеет положительно заряженные боковые цепи
остаток. Притяжение этих зарядов к отрицательным внутренностям
покоящиеся клетки помогают держать канал закрытым. б) когда мембрана становится
деполяризованные (снаружи отрицательные), чувствительные к напряжению спирали движутся к
внешней поверхности плазматической мембраны, вызывая немедленное конформационное изменение
сегмент ворот, открывающий канал для поступления ионов Na+. (c) В течение
миллисекунды после открытия спирали, чувствительные к напряжению, возвращаются в исходное положение.
положение, и сегмент инактивации канала (фиолетовый) перемещается в открытый
канала, препятствуя дальнейшему движению ионов. Когда мембранный потенциал
перевернутый так, что внутренняя часть снова отрицательная, ворота перемещаются обратно в
положение блокировки (не показано). Через 1 2 мс канал деактивируется.
сегмент вытесняется из отверстия канала, и белок возвращается к
закрытое состояние покоя (а), где он может быть снова открыт путем деполяризации.