
Контроль грунта и асфальта
Прибор стандартного уплотнения грунта ПСУ-МГ4 предназначен для уплотнения грунта путем послойного (в три слоя) уплотнения образца грунта с постоянной работой уплотнения, при испытании грунтов по ГОСТ 22733-2002 «Грунты. Метод лабораторного определения максимальной плотности». Метод заключается в установлении зависимости плотности сухого грунта от его влажности при уплотнении образцов грунта с постоянной работой уплотнения и последовательным увеличением влажности.
Принцип работы прибора — автоматический подъем и сброс грузов на уплотняемый материал с постоянной высоты, с автоматическим отсчетом количества ударов. Прибор ПСУ-МГ4 обеспечивает одновременное испытание двух образцов грунта в полуавтоматическом режиме. Оснащен механизмом подъема и сброса грузов, счетчиком числа ударов, клавиатурой для ввода исходных данных и цифровым дисплеем.
Подробнее…
Измеритель степени пучинистости грунта УПГ-МГ4. 01/Н Грунт предназначен для измерений температуры и перемещения поверхности образца грунта при его промораживании в заданном температурном режиме. Принцип действия измерителя основан на изменении сопротивления потенциометрического датчика перемещения в зависимости от вертикальной деформации образца грунта при его промораживании. Для измерения температуры образца грунта применяются терморезисторы, электрическое сопротивления которых изменяется в зависимости от их температуры. Сигналы с датчиков температуры и перемещения регистрируются блоком управления, обрабатываются, и результаты измерений в единицах длины и температуры отображаются на дисплее.
01/Н Грунт предназначен для измерений температуры и перемещения поверхности образца грунта при его промораживании в заданном температурном режиме. Принцип действия измерителя основан на изменении сопротивления потенциометрического датчика перемещения в зависимости от вертикальной деформации образца грунта при его промораживании. Для измерения температуры образца грунта применяются терморезисторы, электрическое сопротивления которых изменяется в зависимости от их температуры. Сигналы с датчиков температуры и перемещения регистрируются блоком управления, обрабатываются, и результаты измерений в единицах длины и температуры отображаются на дисплее.
Подробнее…
Пенетрометр автоматический для нефтебитумов АПН-360МГ4 предназначен для определения пенетрации (глубины проникания иглы) в испытуемый образец при заданных нагрузке и температуре по ГОСТ 33136 и ГОСТ 11501, в соответствии с МИ2418. Прибор АПН-360МГ4 при этом обеспечивает автоматическую регистрацию глубины проникания иглы и времени испытаний. Графический дисплей отображает процесс испытаний. Остановка процесса испытаний — автоматическая, с занесением результата в архив прибора. Прибор оснащен пузырьковым уровнем, регулировочными ножками, подъемным столом, подсветкой иглы и имеет функцию передачи данных на ПК.
Прибор АПН-360МГ4 при этом обеспечивает автоматическую регистрацию глубины проникания иглы и времени испытаний. Графический дисплей отображает процесс испытаний. Остановка процесса испытаний — автоматическая, с занесением результата в архив прибора. Прибор оснащен пузырьковым уровнем, регулировочными ножками, подъемным столом, подсветкой иглы и имеет функцию передачи данных на ПК.
Подробнее…
Толщиномер дорожного покрытия StratoTest 4100 немецкой компании ElektroPhysik предназначен для неразрушающего измерения толщины любых неметаллических слоев дорожного полотна, таких как асфальт, песок, щебень, гравий, шлак, бетон и т. д. Принцип работы – вихретоковый. Диапазон измерений от 0 до 40 см., со специальным датчиком до 80 см.
Для использования толщиномера под выбранными участками будущего полотна подкладывается алюминиевая фольга, которая служит отражателем для датчика, преобразующего сигнал в значение толщины.
Подробнее…
Статический пенетрометр ПСГ-МГ4 предназначен для оперативного контроля качества уплотнения и прочностных характеристик грунта, таких как угол внутреннего трения, удельное сцепление и модуль упругости. Диапазон измерения силы пенетрации 100 — 950 Н. Относительная погрешность измерения ± 1%. Область применения – контроль качества уплотнения грунтов и оснований при строительстве дорог, мостов, опор, железнодорожного полотна, фундаментов, каналов, траншей.
Принцип действия пенетрометра ПСГ-МГ4 заключается в преобразовании деформации упругого элемента тензометрического датчика силы, вызванной действием приложенной силы, в электрический сигнал. Электрический сигнал регистрируется электронным блоком, обрабатывается, и результаты измерений в единицах силы отображаются на дисплее.
Электрический сигнал регистрируется электронным блоком, обрабатывается, и результаты измерений в единицах силы отображаются на дисплее.
Подробнее…
Плотномер асфальтобетона ПА-МГ4 предназначен для измерения плотности, коэффициента уплотнения и температуры асфальтобетонных покрытий. Диапазон измерений плотности от 1900 до 2700 кг/м3. Диапазон измерений температуры от минус 10 до 140ºС. Область применения – оперативный контроль плотности и равномерности уплотнения асфальтобетонных покрытий на предприятиях дорожного строительства и в испытательных лабораториях.
Принцип работы плотномера ПА-МГ4 основан на корреляционной зависимости между амплитудой высокочастотного электромагнитного сигнала и плотностью асфальта / бетона. При взаимодействии с измеряемым материалом преобразователь вырабатывает сигнал, который регистрируется электронным блоком и преобразуется в значение плотности. Предусмотрены три режима измерения: непрерывный; одиночный; с усреднением результата по серии измерений.
Подробнее…
Динамический плотномер грунтов ПДУ-МГ4 «Удар» и его модификация ПДУ-МГ4.01 предназначены для измерения модуля упругости грунтов и дорог на основе прямых измерений амплитуды перемещения штампа и действующей на него ударной силы. Область применения – контроль качества уплотнения грунтов и оснований при строительстве дорог, мостов, опор, железнодорожного полотна, фундаментов, каналов и траншей на стройплощадках и в лабораторных условиях. Конструктивно плотномеры ПДУ-МГ4 состоят из нагрузочной плиты, с закрепленными на ней тензодатчиком силы, акселерометром и упругим элементом, штанги с грузом и электронного блока.
Принцип действия плотномера ПДУ-МГ4 основан на преобразовании деформации упругих элементов тензометрического датчика, возникающей под действием силы, в аналоговый электрический сигнал, изменяющийся пропорционально приложенной силе. Для преобразования величины ускорения штампа в перемещение применяется цифровой интегратор.
Подробнее…
| 5.1 | ГОСТ 33133-2014 | Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Технические требования. Приказ Федерального агентства по техническому регулированию и метрологии от 29 мая 2015 г. N 520-ст |
| 5.2 | ГОСТ 33134-2014 | Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Определение индекса пенетрации. Приказ Федерального агентства по техническому регулированию и метрологии от 29 мая 2015 г. N 521-ст |
| 5.3 | ГОСТ 11508-74 | Битумы нефтяные. Методы определения сцепления битума с мрамором и песком. Постановление Государственного комитета стандартов Совета Министров СССР от 4 февраля 1974 г. N 336 Методы определения сцепления битума с мрамором и песком. Постановление Государственного комитета стандартов Совета Министров СССР от 4 февраля 1974 г. N 336
|
| 5.4 | ГОСТ 33136-2014 | Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Метод определения глубины проникания иглы. Приказ Федерального агентства по техническому регулированию и метрологии от 21 июля 2015 г. N 917-ст |
| 5.5 | ГОСТ 33137-2014 | Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Метод определения динамической вязкости ротационным вискозиметром. Приказ Федерального агентства по техническому регулированию и метрологии от 29 мая 2015 г. N 523-ст |
| 5.6 | ГОСТ 33138-2014 | Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Метод определения растяжимости. Приказ Федерального агентства по техническому регулированию и метрологии от 29 мая 2015 г. |
| 5.8 | ГОСТ 33141-2014 | Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Метод определения температур вспышки. Метод с применением открытого тигля Кливленда. Приказ Федерального агентства по техническому регулированию и метрологии от 29 мая 2015 г. N 527-ст |
| 5.9 | ГОСТ 33142-2014 | Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Метод определения температуры размягчения. Метод «Кольцо и Шар». Приказ Федерального агентства по техническому регулированию и метрологии от 29 мая 2015 г. N 528-ст |
| 5.15 | ГОСТ Р 52128-2003 | Эмульсии битумные дорожные. Технические условия. Постановление Госстроя России от 27 июня 2003 г. N 117 |
| 5.27 | ГОСТ 2517-2012 | Нефть и нефтепродукты. Методы отбора проб. Приказ Федерального агентства по техническому регулированию и метрологии от 29 ноября 2012 г. N 1448-ст N 1448-ст
|
| 5.28 | ГОСТ 22245-90 | Битумы нефтяные дорожные вязкие. Технические условия. Постановление Государственного комитета СССР по стандартам от 12.02.90 N 191 |
| 5.29 | ГОСТ 11501-78 | Битумы нефтяные. Метод определения глубины проникания иглы. Постановление Государственного Комитета СССР по стандартам от 06.09.78 N 2457 |
| 5.30 | ГОСТ 11505-75 | Битумы нефтяные. Метод определения растяжимости. Постановление Государственного комитета стандартов Совета Министров СССР от 29.12.75 N 4072 |
| 5.31 | ГОСТ 11506-73 | Битумы нефтяные. Метод определения температуры размягчения по кольцу и шару. Постановление Государственного комитета СССР по стандартам от 18.07.73 N 1753 |
| 5.32 | ГОСТ 11507-78 | Битумы нефтяные. Метод определения температуры хрупкости по Фраасу. Постановление Государственного комитета СССР по стандартам от 11 декабря 1978 г. N 3281 Постановление Государственного комитета СССР по стандартам от 11 декабря 1978 г. N 3281
|
| 5.33 | ГОСТ 4333-2014 | Нефтепродукты. Методы определения температур вспышки и воспламенения в открытом тигле. Приказ Федерального агентства по техническому регулированию и метрологии от 29 мая 2015 г. N 470-ст |
| 6 |
Цемент | |
| 6.1 | ГОСТ 33174-2014 | Дороги автомобильные общего пользования. Цемент. Технические требования. Приказ Федерального агентства по техническому регулированию и метрологии от 26 марта 2015 г. N 179-ст |
| 6.2 | ГОСТ Р 55224-2012 | Цементы для транспортного строительства. Технические условия. Приказ Федерального агентства по техническому регулированию и метрологии от 27 ноября 2012 г. N 1248-ст |
| 6.3 | ГОСТ 31108-2003 | Цементы общестроительные. Технические условия. Постановление Госстроя России от 21 июня 2003 г. N 93 Технические условия. Постановление Госстроя России от 21 июня 2003 г. N 93
|
| 6.4 | ГОСТ 30515-2013 | Цементы. Общие технические условия. Приказ Федерального агентства по техническому регулированию и метрологии от 11 июня 2014 г. N 654-ст |
| 6.5 | ГОСТ 10178-85 | Портландцемент и шлакопортландцемент. Технические условия. Постановление Государственного комитета СССР по делам строительства от 10.07.85 N 116 |
| 7 |
Грунты | |
| 7.1 | ГОСТ 25100-2011 | Грунты. Классификация. Приказ Федерального агентства по техническому регулированию и метрологии от 12 июля 2012 г. N 190-ст |
| 7.2 | ГОСТ 12071-2014 | Грунты. Отбор, упаковка, транспортирование и хранение образцов. Приказ Федерального агентства по техническому регулированию и метрологии от 12 декабря 2014 г. N 2023-ст N 2023-ст
|
| 7.3 | ГОСТ 30672-2012 | Грунты. Полевые испытания. Общие положения. Приказ Федерального агентства по техническому регулированию и метрологии от 29 октября 2012 г. N 594-ст |
| 7.4 | ГОСТ 5180-2015 | Грунты. Методы лабораторного определения физических характеристик. Приказ Федерального агентства по техническому регулированию и метрологии от 3 ноября 2015 г. N 1694-ст |
| 7.5 | ГОСТ 22733-2002 | Грунты. Метод лабораторного определения максимальной плотности. Постановление Госстроя России от 27 декабря 2002 г. N 170 |
| 7.6 | ГОСТ 25584-90 | Грунты. Методы лабораторного определения коэффициента фильтрации. Постановление Государственного комитета СССР от 04.04.90 N 32 |
| 8 |
Рекомендуемые страницы: |
|
Битум БНД 70/100
Битум нефтяной дорожный вязкий БНД 70/100 изготавливается согласно ГОСТ 33133-2014 Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Технические требования.
Битумы нефтяные дорожные вязкие. Технические требования.
Применяется в качестве вяжущего материала при строительстве и ремонте дорожных покрытий и оснований, а также в качестве основы для производства модифицированных битумов и битумных эмульсий.
Марка БНД 70/100 занимает промежуточную позицию, между более жидким БНД 90/130 и более вязким БНД 60/90.
Физико-химические показатели БНД 70/100:
| № п/п | Наименование показателя, единица измерения | Метод испытания | Нормы по ГОСТ 33133-2014 БНД 70/100 |
| 1 | Глубина проникания иглы 0,1мм: при 25°С | ГОСТ 33136-2014 | 71-100 |
| 2 | Температура размягчения по кольцу и шару, оС | ГОСТ 33142-2014 | не ниже 47 |
| 3 | Растяжимость, см, при 0 °С | ГОСТ 33138-2014 | не менее 3,7 |
| 4 | Температура хрупкости, °С | ГОСТ 33143-2014 | не выше -18 |
| 5 | Температура вспышки, °С | ГОСТ 33141-2014 | не ниже 230 |
| 6 | Изменение массы образца после старения, % | ГОСТ 33140-2014 | не более 0,6 |
| 7 | Изменение температуры размягчения после старения, 0С | ГОСТ 33140-2014 ГОСТ 33142-2014 | не более 7,0 |
ООО «ПСП «Карьер Октябрьское» 8(496)616-40-44
Аттестация аппарата ЛинтеЛ ДБ–20-150 для определения растяжимости нефтяных битумов
ГОСТ 11505-75, ГОСТ 33136-2014, ГОСТ 33138-2014, ASTM D-113-99, ASTM D-6084-04, ГОСТ Р 52056-2003, ГОСТ 11262-80. Аттестация аппарата ЛинтеЛ ДБ–20-150 для определения растяжимости нефтяных битумов для определения растяжимости нефтяных битумов осуществляется в аккредитованной лаборатории РЦСМ и занимает от 1 до 5 дней.
Аттестация аппарата ЛинтеЛ ДБ–20-150 для определения растяжимости нефтяных битумов для определения растяжимости нефтяных битумов осуществляется в аккредитованной лаборатории РЦСМ и занимает от 1 до 5 дней.
Аттестация аппарата ЛинтеЛ ДБ–20-150 для определения растяжимости нефтяных битумов
Аппарат предназначен для определения растяжимости нефтебитумов и других пластических материалов. Температура регулируется с помощью термостата ТС-20 или термокриостата ТКС-20 автоматически.
Автоматические функции аппарата:- измерение и индикацию температуры теплоносителя в зоне испытаний;
- растяжение одновременно трёх образцов с заданной скоростью с фиксацией и запоминанием значения растяжимости и номера пробы;
- перемешивание теплоносителя для равномерной температуры в зоне испытания;
- проведение испытания на релаксацию для одного образца в следующей последовательности:
- 1. растяжение до заданной длины или минимальной силы с постоянной скоростью;
2. выдержка при заданном времени;
выдержка при заданном времени;
3. сжатие нити с заданной скоростью и поддержание минимальной силы натяжения нити после растяжения в заданное время или до падения скорости сжатия нити ниже заданной. - запоминание, хранение и выдача до 30 последних графиков: растяжения, выдержки, сжатия;
- запоминание и хранение до 100 последних результатов испытаний с выдачей на дисплей и компьютер: растяжимости, длины после сжатия, эластичности и остаточного расширения, температуры в ходе испытания, двух максимальных значений силы натяжения нити и соответствующей длины нити в ходе испытания, названия пробы, имени лаборанта и времени проведения испытания;
- остановка процесса растяжения по окончании испытания;
- автоматическая передача результатов по радиоканалу на ПК через «ПБИ ЛинтеЛ-Линк» в базу данных системы «ЛинтеЛ-ЛИС».
 выдержка при заданном времени;
выдержка при заданном времени;ВНИМАНИЕ: «ЛинтеЛ-Линк» и «ЛинтеЛ-ЛИС» приобретаются отдельно.
RussianGost | Официальная нормативная библиотека — ГОСТ 33133-2014
Товар содержится в следующих классификаторах:
Конструкция (макс. ) »
Стандарты »
Прочие государственные стандарты, применяемые в строительстве »
93 Гражданское строительство »
) »
Стандарты »
Прочие государственные стандарты, применяемые в строительстве »
93 Гражданское строительство »
Доказательная база (ТР ТС, Технический регламент Таможенного союза) » 014/2011 ТР КС. Безопасность дорожного движения » Нормы и стандарты (к ТР ТС 014/2011) »
Классификатор ISO » 93 ГРАЖДАНСКОЕ СТРОИТЕЛЬСТВО » 93.080 Дорожное строительство » 93.080.20 Дорожно-строительные материалы »
Национальные стандарты » 93 ГРАЖДАНСКОЕ СТРОИТЕЛЬСТВО » 93.080 Дорожное строительство » 93.080.20 Дорожно-строительные материалы »
Ссылки на документы:
ГОСТ 12.1.004-91 — Пожарная безопасность. Общие требования
ГОСТ 12.1.005-88 — Общие санитарные требования к воздуху рабочей зоны
.
ГОСТ 12.1.007-76: Вредные вещества. Классификация и общие требования безопасности
ГОСТ 12.1.014-84 — Воздух рабочей зоны
.ГОСТ 12.1.044-89 — Пожаровзрывоопасность веществ и материалов. Номенклатура показателей и методы их определения
ГОСТ 1510-84 — Нефть и нефтепродукты. Маркировка, упаковка, транспортировка и хранение
ГОСТ 2517-2012 — Нефть и нефтепродукты. Методы отбора проб
ГОСТ 33134-2014 — Дороги автомобильные общего пользования.Битумы нефтяные дорожные вязкие. Определение индекса проникновения
ГОСТ 33135-2014 — Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Метод определения растворимости
ГОСТ 33136-2014 — Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Метод определения глубины проникновения иглы пенетрометра
ГОСТ 33137-2014 — Дороги общего пользования. Битум нефтяной вязкий для дорожного строительства. Метод определения динамической вязкости ротационным вискозиметром
ГОСТ 33138-2014 — Дороги общего пользования. Битум нефтяной вязкий для дорожного строительства. Метод определения расширяемости
Битум нефтяной вязкий для дорожного строительства. Метод определения расширяемости
ГОСТ 33139-2014 — Дороги автомобильные общего пользования. Битум нефтяной дорожный вязкий. Метод определения содержания твердых парафинов
ГОСТ 33140-2014 — Дороги автомобильные общего пользования. Битум нефтяной дорожный вязкий. Метод определения старения под воздействием высокой температуры и воздуха (метод RTFOT)
ГОСТ 33141-2014 — Дороги автомобильные общего пользования.Битумы нефтяные дорожные вязкие. Определение точек возгорания. Метод открытого стакана Кливленда
ГОСТ 33142-2014 — Дороги общего пользования. Битум нефтяной вязкий для дорожного строительства. Метод определения точки размягчения. Метод «кольцо и мяч»
ГОСТ 33143-2014 — Дороги общего пользования. Битум нефтяной вязкий для дорожного строительства. Метод определения температуры разрушения по методу Фрааса
Ссылка на документ:
ГОСТ 33134-2014 — Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Определение индекса проникновения
Битумы нефтяные дорожные вязкие. Определение индекса проникновения
ГОСТ 33135-2014 — Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Метод определения растворимости
ГОСТ 33136-2014 — Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Метод определения глубины проникновения иглы пенетрометра
ГОСТ 33137-2014 — Дороги общего пользования. Битум нефтяной вязкий для дорожного строительства. Метод определения динамической вязкости ротационным вискозиметром
ГОСТ 33138-2014 — Дороги общего пользования.Битум нефтяной вязкий для дорожного строительства. Метод определения расширяемости
ГОСТ 33139-2014 — Дороги автомобильные общего пользования. Битум нефтяной дорожный вязкий. Метод определения содержания твердых парафинов
ГОСТ 33140-2014 — Дороги автомобильные общего пользования. Битум нефтяной дорожный вязкий. Метод определения старения под воздействием высокой температуры и воздуха (метод RTFOT)
ГОСТ 33141-2014 — Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Определение точек возгорания. Метод открытого стакана Кливленда
Битумы нефтяные дорожные вязкие. Определение точек возгорания. Метод открытого стакана Кливленда
ГОСТ 33142-2014 — Дороги общего пользования. Битум нефтяной вязкий для дорожного строительства. Метод определения точки размягчения. Метод «кольцо и мяч»
ГОСТ 33143-2014 — Дороги общего пользования. Битум нефтяной вязкий для дорожного строительства. Метод определения температуры разрушения по методу Фрааса
ODM 218.2.087-2017: Рекомендации по проектированию и устройству водопропускных труб из спирально-волнистых металлических гофрированных труб
ODM 218.3.060-2015: Методические указания по ремонту покрытий из цементобетона, покрытых асфальтобетонными покрытиями, на дорогах общего пользования
ОДМ 218.3.063-2016: Рекомендации по выбору битумных вяжущих на основе определения их реологических свойств для повышения стойкости к образованию асфальтобетона
ODM 218.3.082-2016: Методические указания по назначению технологии и периодичности работ по устройству слоев износа и защитных слоев дорожных покрытий
ODM 218. 3.087-2017: Рекомендации по применению асфальтобетонных смесей на основе металлургических шлаковых материалов для условий ЦФО
3.087-2017: Рекомендации по применению асфальтобетонных смесей на основе металлургических шлаковых материалов для условий ЦФО
ODM 218.3.102-2017: Методические указания по устройству асфальтового покрытия в сложных погодных условиях
ОДМ 218.4.038-2017: Методические указания по приемке покрытий из плотных асфальтобетонных смесей, рассчитанных объемным методом
ПНСТ 183-2016 — Дороги автомобильные общего пользования. Битумно-каменно-мастичные смеси и каменно-мастичный асфальт.Технические характеристики
ПНСТ 184-2016 — Дороги автомобильные общего пользования. Дорожные горячие асфальтобетонные смеси и асфальтобетон. Технические характеристики
ПНСТ 191-2017 — Смеси сероасфальтобетонные и сероасфальтобетонные. Технические характеристики
ПНСТ 192-2017 — Смесь щебеночно-мастичного сероасфальта и каменно-мастичного сероасфальта. Технические характеристики
ПНСТ 265-2018 — Дороги автомобильные общего пользования. Конструкция гибкого покрытия
ПНСТ 266-2018 — Смесь серно-асфальтобетонная литая и асфальтобетонный литой. Технические характеристики
Технические характеристики
ГОСТ Р 54401-2020 — Дороги автомобильные общего пользования. Горячие асфальтобетонные и горячие асфальтобетонные смеси. Технические характеристики
ГОСТ Р 58397-2019 — Дороги автомобильные общего пользования. Правила изготовления работ. Оценка соответствия
ГОСТ Р 58406.2-2020 — Дороги автомобильные общего пользования. Горячие смеси, асфальтобетон и асфальтобетон. Технические характеристики
ГОСТ Р 58818-2020 — Дороги автомобильные с малой интенсивностью движения. Дизайн, конструкция и расчет
ГОСТ Р 58829-2020 — Дороги автомобильные общего пользования.Битумы нефтяные дорожные вязкие. Правила выбора марок в зависимости от прогнозируемых транспортных нагрузок и климатических условий эксплуатации на основе дополнительных показателей
ГОСТ Р 58831-2020 — Дороги автомобильные общего пользования. Покрытия асфальтобетонные. Общие правила использования прибора в сложных погодных условиях
ГОСТ Р 59290-2021 — Дороги автомобильные общего пользования. Требования к ведению входящего и оперативного контроля
Требования к ведению входящего и оперативного контроля
ODM 218.2.104-2019: Альбом типовых конструкций нежестких покрытий в различных климатических зонах
ODM 218.3.064-2019: Методические указания по оценке эффективности адгезионных добавок в составе асфальтобетона
.ОДМ 218.3.084-2020: Рекомендации по приготовлению и применению органо-минеральных смесей при устройстве конструкционных слоев дорожных покрытий капитального и легкого типов
ODM 218.3.096-2017: Руководство по объемному расчету асфальтобетонных смесей по методологии Маршалла
ODM 218.4.009-2017: Методические рекомендации по технологии пылеулавливания дорог с переходным типом покрытия битумной эмульсией
ODM 218.8.009-2017: Методические рекомендации по технологии пылеулавливания дорог с переходным типом покрытия битумной эмульсией
ПНСТ 183-2019 — Дороги автомобильные общего пользования. Смешивает асфальтобетон дорожный и асфальтобетонный щебень и мастику. Технические характеристики
Технические характеристики
ПНСТ 184-2019 — Дороги автомобильные общего пользования. Смеси дорожный асфальтобетон и асфальтобетон. Технические характеристики
ПНСТ 325-2019 — Дороги автомобильные общего пользования. Смеси щебня, гравия и песка, обработанные органическими вяжущими.Технические характеристики
ПНСТ 362-2019 — Дороги автомобильные общего пользования. Холодные асфальтовые смеси и асфальтобетонные смеси. Технические условия
ПНСТ 371-2019 — Дороги общего пользования с низкой плотностью движения. Дорожная одежда. Конструкция и расчет
ПНСТ 390-2020 — Дороги автомобильные общего пользования. Нежесткая дорожная одежда. Типовые исполнения
Клиентов, которые просматривали этот товар, также просматривали:
|
 Оценки
Оценки Определение плотности жидкостей и твердых тел
Определение плотности жидкостей и твердых тел Методы испытаний
Методы испытаний Общие требования безопасности
Общие требования безопасностиВАШ ЗАКАЗ ПРОСТО!
Русский Гост.com — ведущая в отрасли компания со строгими стандартами контроля качества, и наша приверженность точности, надежности и аккуратности является одной из причин, почему некоторые из крупнейших мировых компаний доверяют нам обеспечение своей национальной нормативно-правовой базы и перевод критических, сложных и конфиденциальная информация.
Наша нишевая специализация — локализация национальных нормативных баз данных, включающих: технические нормы, стандарты и правила; государственные законы, кодексы и постановления; а также кодексы, требования и инструкции агентств РФ.
У нас есть база данных, содержащая более 220 000 нормативных документов на английском и других языках для следующих 12 стран: Армения, Азербайджан, Беларусь, Казахстан, Кыргызстан, Молдова, Монголия, Россия, Таджикистан, Туркменистан, Украина и Узбекистан.
Размещение заказа
Выберите выбранный вами документ, перейдите на «страницу оформления заказа» и выберите желаемую форму оплаты. Мы принимаем все основные кредитные карты и банковские переводы.Мы также принимаем PayPal и Google Checkout для вашего удобства. Свяжитесь с нами для любых дополнительных договоренностей (договорные соглашения, заказ на поставку и т. Д.).
После размещения заказа он будет проверен и обработан в течение нескольких часов, но в редких случаях — максимум 24 часа.
Документ / веб-ссылка для товаров на складе будет отправлена вам по электронной почте, чтобы вы могли загрузить и сохранить ее для своих записей.
Если товары отсутствуют на складе (поставка сторонних поставщиков), вы будете уведомлены о том, для каких товаров потребуется дополнительное время.Обычно мы поставляем такие товары менее чем за три дня.
Как только заказ будет размещен, вы получите квитанцию / счет, который можно будет заполнить для отчетности и бухгалтерского учета. Эту квитанцию можно легко сохранить и распечатать для ваших записей.
Гарантия лучшего качества и подлинности вашего заказа
Ваш заказ предоставляется в электронном формате (обычно это Adobe Acrobat или MS Word).
Мы всегда гарантируем лучшее качество всей нашей продукции.Если по какой-либо причине вы не удовлетворены, мы можем провести совершенно БЕСПЛАТНУЮ ревизию и редактирование приобретенных вами продуктов. Кроме того, мы предоставляем БЕСПЛАТНЫЕ обновления нормативных требований, если, например, документ имеет более новую версию на дату покупки.
Гарантируем подлинность. Каждый документ на английском языке сверяется с оригинальной и официальной версией. Мы используем только официальные нормативные источники, чтобы убедиться, что у вас самая последняя версия документа, причем все из надежных официальных источников.
Экспериментальное исследование неокисленного и окисленного битума, полученного из тяжелой нефти
Наваз, С.М. Н. и Алви, С. Энергетическая безопасность для социально-экономической и экологической устойчивости в Пакистане. Heliyon 4 (10), e00854 (2018).
Артикул Google Scholar
Карре, Д. Ле Битум, Un Matériau Pour Le Développement Durable. RGRA-Revue Generale des Routes et des Aerodromes 2010 , No.883, 47
Де Кастро, К., Мигель, Л. Дж. И Медиавилла, М. Роль нетрадиционного масла в ослаблении пика масла. Энергетическая политика 37 (5), 1825–1833 (2009).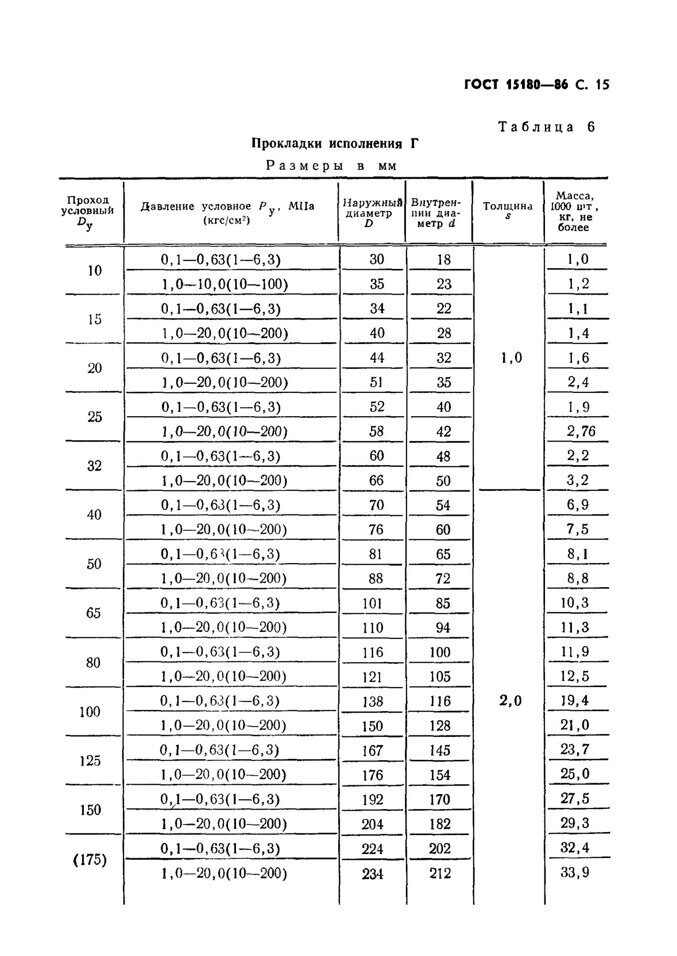
Артикул Google Scholar
Джаякоди, С., Галлаж, К. и Рамануджам, Дж. Рабочие характеристики переработанного заполнителя бетона в качестве несвязанного материала дорожного покрытия. Heliyon 5 (9), e02494 (2019).
CAS PubMed PubMed Central Статья Google Scholar
Sun, F. et al. Характеристики потока и теплопередачи перегретого пара в морских скважинах и анализ характеристик перегретого пара. Comput. Chem. Англ. 100 , 80–93 (2017).
CAS Статья Google Scholar
Sun, F. et al. Анализ эффективности закачки перегретого пара для извлечения тяжелой нефти и моделирование тепловой эффективности ствола скважины. Energy 125 , 795–804 (2017).
Артикул Google Scholar

Джимасбе, Р., Аль-мунтасер, А. А., Сувайд, М. А., В и арфоломеев, М. А. Сравнение обогащения тяжелой нефти и остатков вакуумной перегонки водой в сверхкритическом состоянии. В Серия конференций IOP: Науки о Земле и окружающей среде ; IOP Publishing, 2019; Vol.282, p. 12044
Djimasbe, R. et al. Исследование технологии производства модифицированных серных битумных вяжущих. Бык. Воронежский гос. Ун-т. Англ. Technol. 80 (2), 76 (2018).
Google Scholar
Sun, F., Yao, Y. & Li, X. Характеристики тепломассопереноса перегретого пара в сочетании с неконденсирующимися газами в горизонтальных скважинах с многоточечной закачкой. Энергетика 143 , 995–1005 (2018).
Артикул Google Scholar
Лю П., Му, З., Ли, В., Ву, Ю. и Ли, X. Новая математическая модель и экспериментальная проверка течения пенистой нефти при разработке пластов тяжелой нефти. Sci. Отчет 7 (1), 1–13 (2017).
Sci. Отчет 7 (1), 1–13 (2017).
ADS Статья Google Scholar
Chengzao, J., Zheng, M. & Zhang, Y. Нетрадиционные углеводородные ресурсы в Китае и перспективы разведки и разработки. Pet. Explor. Dev. 39 (2), 139–146 (2012).
Артикул Google Scholar
Аллен Р. Г., Литтл Д. Н., Бхасин А. и Гловер К. Дж. Влияние химического состава на микроструктуру асфальта и их связь с характеристиками дорожного покрытия. Внутр. J. Pavement Eng. 15 (1), 9–22 (2014).
CAS Статья Google Scholar
Абаев Г. Н., Кушнир Ю. В., Дубровский А. В., Михайлова О. Н., Андреева Р. А., Димуду И. А., Клюев А. И. Развитие методов испытаний для перегонки нефтепродуктов. Ind. Serv. 2013 48 (3)
Саней Х. Генезис твердых битумов. Sci. Отчет 10 (1), 1–10 (2020).
Sci. Отчет 10 (1), 1–10 (2020).
ADS Статья Google Scholar
Фулчер К., Стейси Р. и Спенсер Н. Битум из мертвого моря в раннем железном веке Нубии. Sci. Отчет 10 (1), 1–12 (2020).
Артикул Google Scholar
Рудык С. Взаимосвязь между SARA фракциями обычной нефти и тяжелой нефти. Natl. Остатки битума. Топливо 216 , 330–340 (2018).
CAS Google Scholar
Туробова М.А., Данилов В.Э., Айзенштадт А.М. Использование отходов деревообрабатывающей промышленности для модификации битума. В Серия конференций IOP: Материаловедение и инженерия ; IOP Publishing, 2020; Vol. 945, p 12061
Морозов В.А., Старов Д.С., Шахова Н.М., Колобков В.С. Производство дорожных асфальтов из нефтей с высоким содержанием парафина. Chem. Technol. Топливные масла 40 (6), 382–388 (2004).
Chem. Technol. Топливные масла 40 (6), 382–388 (2004).
CAS Статья Google Scholar
Чжан, Х., Чжу, К., Ю, Дж., Тан, Б. и Ши, С. Влияние нано-оксида цинка на свойства ультрафиолетового старения битума со степенью пенетрации 60/80. Mater. Struct. 48 (10), 3249–3257 (2015).
CAS Статья Google Scholar
Курлыкина А., Денисов В., Кузнецов Д., Лукаш Е. А. Щебеночно-мастичный асфальтобетон с использованием серововмещающих технологий. Вестник Белгородского государственного технологического университета.Шухов В.Г. 2020 , № 1
Абдуллин А.И., Идрисов М.Р., Емельянычева Е.П. Повышение термоокислительной устойчивости нефтяных битумов с использованием технологии «пероксидирование – разбавление» и введение антиоксидантных присадок. Pet. Sci. Technol. 35 (18), 1859–1865 (2017).
CAS Статья Google Scholar
Соенен, Х., Лу, X. и Лаукканен, О.-V. Окисление битума: молекулярная характеристика и влияние на реологические свойства. Rheol. Acta 55 (4), 315–326 (2016).
CAS Статья Google Scholar
Чаала А., Чиочина О. Г. и Рой К. Вакуумный пиролиз остатков автомобильного измельчителя: использование пиролитического масла в качестве модификатора дорожного битума. Resour. Консерв. Recycl. 26 (3–4), 155–172 (1999).
Артикул Google Scholar
Чаала А., Рой К. и Айт-Кади А. Реологические свойства битума, модифицированного пиролитической сажей. Топливо 75 (13), 1575–1583 (1996).
CAS Статья Google Scholar
Shah, A. et al. Обзор новых технологий добычи и обогащения тяжелой нефти и битума. Energy Environ. Sci. 3 (6), 700–714 (2010).
Energy Environ. Sci. 3 (6), 700–714 (2010).
CAS Статья Google Scholar
Лодерер, К., Партл, М. Н. и Пуликакос, Л. Д. Влияние технологии производства резиновой крошки на характеристики модифицированного битума. Констр. Строить. Матер. 191 , 1159–1171 (2018).
Артикул Google Scholar
Мардупенко А., Григоров А., Синкевич И., Тульская А. Технология производства модифицированного битума для дорожного строительства. 2019
Cheraghian, G. & Wistuba, M.P. Исследование ультрафиолетового старения битума, модифицированного композитом из глины и наночастиц коллоидного диоксида кремния. Sci. Отчет 10 (1), 1–17 (2020).
Артикул Google Scholar
Кок М.В. Термические исследования сейитомерного горючего сланца. Термохим. Acta 369 (1–2), 149–155 (2001).
Термохим. Acta 369 (1–2), 149–155 (2001).
CAS Статья Google Scholar
Ришвана С.С., Махендран А. и Виджаякумар С.Т. Исследования структурно различных бензоксазинов на основе дифенолов и диаминов: кинетика термического разложения и исследования TG-FTIR. Термохим. Acta 618 , 74–87 (2015).
Артикул Google Scholar
Саванчук Р.А. Анализ влияния группового состава нефтепродуктов на показатели качества дорожных битумов. Альманах World Sci. 6 , 20–23 (2019).
Google Scholar
Абдуллин А., Идрисов М., Ганиева Т., Емельянычева Е. Водно-битумные эмульсии ; Литров, 2017
Волков В.Ю., Аль-Мунтасер А.А., Варфоломеев М.А., Хасанова Н.М., Сахаров, Б. В., Сувайд, М. А., Джимасбе, Р., Галеев, Р. И., и Нургалиев, Д. К. Низкополевая ЯМР-релаксометрия как быстрый и простой метод определения SARA-состава сырой нефти на месте. J. Pet. Sci. Англ. 196 , 107990
К. Низкополевая ЯМР-релаксометрия как быстрый и простой метод определения SARA-состава сырой нефти на месте. J. Pet. Sci. Англ. 196 , 107990
Ядыкина В.В., Акимов А.Е., Траутвайн А.И., Холопов В.С. Влияние понижающей температуры добавки ДАД-ТА на физико-механические свойства битума и уплотнение асфальтобетона. В Серия конференций IOP: Материаловедение и инженерия ; IOP Publishing, 2018; Vol.327, p 32006.
Мошреф Х.С., Кутьин Ю.А., Теляшев Е.Г. Нефтяные дорожные битумные материалы. Стандарты, качество, технологии, перспективы. Нефтегазовый бизнес 2012 , № 6
Кемалов А.Ф., Кемалов Р.А., Абдрафикова И.М., Фахретдинов П.С., Валиев Д.З. Полифункциональные модификаторы битумов и битумных материалов с высокими характеристиками. Достижения в области материаловедения и инженерии 2018 , 2018
Pereira, L. et al. Экспериментальное исследование влияния наполнителя на пластичность наполнительно-битумных мастик. Констр. Строить. Матер. 189 , 1045–1053 (2018).
Констр. Строить. Матер. 189 , 1045–1053 (2018).
Артикул Google Scholar
Чжан, Дж., Эйри, Г. Д. и Гренфелл, Дж. Р. А. Экспериментальная оценка прочности когезионного и адгезионного сцепления и энергии разрушения систем битум-заполнитель. Mater. Struct. 49 (7), 2653–2667 (2016).
CAS Статья Google Scholar
Хамидуллин Р., Ковальчук Д., Галиуллин Е. А. Экспериментальные исследования перегонки тяжелых нефтяных остатков в среде инертного газа. Казань Технол. Univ. Бык. 2015 , 18 (20).
Забродин А., Алибеков С. Ю., Забродина Н., Салманов Р., Марьяшев А. Анализ физико-механических свойств мазута М100. Вестник Казанского технологического университета 2013 , 16 (7).
Дедов А.Г., Марченко Д.Ю., Зрелова Л.В., Иванова Е.А. , Санджиева Д.А., Пархоменко А.А., Будинов С.В., Лобакова Е.С., Дольникова Г.А. органических соединений серы в углеводородных средах. Pet. Chem. 2018 , 58 (8), 714–720.
, Санджиева Д.А., Пархоменко А.А., Будинов С.В., Лобакова Е.С., Дольникова Г.А. органических соединений серы в углеводородных средах. Pet. Chem. 2018 , 58 (8), 714–720.
Хомаюни, Ф., Хамиди, А.А. и Ватани, А. Экспериментальное исследование снижения вязкости при транспортировке по трубопроводу тяжелой и сверхтяжелой сырой нефти. Pet. Sci. Technol. 30 (18), 1946–1952 (2012).
CAS Статья Google Scholar
Законы Беларуси | Официальная нормативная библиотека — ГОСТ 17711-93
Продукт содержится в следующих классификаторах:
Стандарты на трубопроводную арматуру (ТПА) » 9.Материалы для трубопроводной арматуры (ТПА) » 9.1 Металлы »
ПромЭксперт »
РАЗДЕЛ I. ТЕХНИЧЕСКИЙ РЕГЛАМЕНТ »
V Тестирование и контроль »
4 Тестирование и контроль продукции »
4. 12 Испытания и контроль продукции металлургической промышленности »
4.12.3 Цветные и драгоценные металлы »
12 Испытания и контроль продукции металлургической промышленности »
4.12.3 Цветные и драгоценные металлы »
Классификатор ISO » 77 МЕТАЛЛУРГИЯ » 77.120 Цветные металлы » 77.120.30 Медь и медные сплавы »
Национальные стандарты » 77 МЕТАЛЛУРГИЯ » 77.120 Цветные металлы » 77.120.30 Медь и медные сплавы »
Национальные стандарты для сомов » Последнее издание » V Металлы и изделия из них » V5 Цветные металлы и их сплавы.Прокат цветных металлов » V51 Цветные металлы, в том числе редкие, и их сплавы »
В качестве замены:
ГОСТ 17711-80 — Сплавы медно-цинковые литейные (латунь)
.Ссылки на документы:
ГОСТ 1497-84 — Металлы. Методы испытания на растяжение
ГОСТ 1652. 13-77 — Сплавы медно-цинковые. Методы определения фосфора
13-77 — Сплавы медно-цинковые. Методы определения фосфора
ГОСТ 1652.1-77 — Сплавы медно-цинковые. Методы определения меди
ГОСТ 17711-80 — Сплавы медно-цинковые литейные (латунь)
.ГОСТ 9012-59 — Металлы.Метод измерения твердости по Бринеллю
Ссылка на документ:
ГОСТ 10007-80 — Политетрафторэтилен. Технические характеристики
ГОСТ 10916-74 — Жидкость силиконовая изоляционная 132-12Д. Технические характеристики
ГОСТ 10944-75 — Арматура регулирующая отопительных установок для систем водяного отопления зданий
.ГОСТ 12.2.052-81 — Система стандартов безопасности труда. Оборудование, работающее с газообразным кислородом. Общие требования безопасности
ГОСТ 12.4.128-83: Защитные каски
ГОСТ 12393-77 — Арматура для воздушных путей электрических железных дорог. Общие технические условия
ГОСТ 14896-84 — Уплотнения резиновые U-образные для гидравлических устройств
. ГОСТ 15151-69 — Машины, приборы и другие промышленные товары для тропиков. Основные Характеристики.
Основные Характеристики.
ГОСТ 15975-70 — Смазка силиконовая типа КБ-3 / 10Э. Технические характеристики
ГОСТ 16336-2013 — Композиции полиэтиленовые для кабельной промышленности. Технические характеристики
ГОСТ 16337-77 — Полиэтилен высокого давления.Технические характеристики
ГОСТ 16338-85 — Полиэтилен низкого давления. Технические характеристики
ГОСТ 1652.10-77 — Сплавы медно-цинковые. Методы определения алюминия
ГОСТ 1652.11-77 — Сплавы медно-цинковые. Методы определения никеля
ГОСТ 1652.12-77 — Сплавы медно-цинковые. Методы определения кремния
ГОСТ 1652.13-77 — Сплавы медно-цинковые. Методы определения фосфора
ГОСТ 1652.1-77 — Сплавы медно-цинковые.Методы определения меди
ГОСТ 1652.2-77 — Сплавы медно-цинковые. Методы определения свинца
ГОСТ 1652.3-77 — Сплавы медно-цинковые. Методы определения железа
ГОСТ 1652.4-77 — Сплавы медно-цинковые. Методы определения марганца
ГОСТ 1652. 5-77 — Сплавы медно-цинковые. Методы определения олова
5-77 — Сплавы медно-цинковые. Методы определения олова
ГОСТ 1652.6-77 — Сплавы медно-цинковые. Методы определения сурьмы
ГОСТ 1652.7-77: Медно-цинковые сплавы. Методы определения висмута
ГОСТ 1652.8-77 — Сплавы медно-цинковые. Методы определения мышьяка
ГОСТ 1652.9-77 — Сплавы медно-цинковые. Методы определения серы
ГОСТ 19681-2016 — Арматура сантехническая и техническая. Общие технические условия
ГОСТ 19681-94 — Приспособления для смешивания и распределения воды санитарно-технические. Общие технические условия
ГОСТ 19689-80 — Обогреватели стекловолоконные тонкослойные
.ГОСТ 2171-90 — Заготовки, изделия, полуфабрикаты и заготовки из цветных металлов и сплавов.Обозначение марки
ГОСТ 21804-94 — Устройства запорные для баллонов с сжиженными углеводородными газами давлением до 1,6 МПа
.ГОСТ 23289-2016 — Комплектующие сантехнические канализационные. Спецификация
ГОСТ 23289-94 — Комплектующие сантехнические канализационные. Технические характеристики
Технические характеристики
ГОСТ 23476-79 — Устройства опоры контактного троллейбуса трамваев и троллейбусов. Общие технические условия
ГОСТ 24301-93 — Прутки и трубы бронзовые и латунные литые.Технические характеристики
ГОСТ 26996-86 — Полипропилен и сополимеры пропилена. Технические характеристики
ГОСТ 30777-2001 — Устройства поворотно-откидные и поворотно-откидные для окон и балконных дверей. Технические характеристики
ГОСТ 30777-2012 — Устройства поворотные, откидные, поворотно-откидные и раздвижные для окон и балконных дверей. Технические характеристики
ГОСТ 30815-2002 — Клапаны автоматические для регулирования температуры в отопительных приборах систем водяного отопления зданий.Общие технические условия
ГОСТ 32803-2014 — Бетон напряженный. Общие технические условия
ГОСТ 33136-2014 — Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Метод определения глубины проникновения иглы пенетрометра
ГОСТ 33142-2014 — Дороги общего пользования. Битум нефтяной вязкий для дорожного строительства. Метод определения точки размягчения. Метод «кольцо и мяч»
Битум нефтяной вязкий для дорожного строительства. Метод определения точки размягчения. Метод «кольцо и мяч»
ГОСТ 33260-2015 — Арматура трубопроводная. Металлы, используемые в производстве трубопроводной арматуры.Основные требования к выбору материалов
ГОСТ 34294-2017 — Арматура трубопроводная криогенная. Общие технические условия
ГОСТ 34347-2017 — Сосуды и аппараты стальные сварные. Общие технические условия
ГОСТ 356-80 — Арматура и детали трубопроводов. Давления условные, испытательные и эксплуатационные. Линии.
ГОСТ 4285-79 — Штуцер продувочный для воздушных рукавов диаметром 19 мм. Технические характеристики
ГОСТ 7171-78 — Смазка бензостойкая. Технические характеристики
ГОСТ 7199-77 — Подшипники резинометаллические судовые.Технические характеристики
ГОСТ 7934.5-74 — Масла и смазки часовые. Метод определения коррозионной активности
ГОСТ 8117-74 — Колокола судовые. Технические характеристики
ГОСТ 982-80 — Масла трансформаторные
.
ГОСТ Р 51177-2017 — Оборудование для воздушных линий. Общие технические требования
ГОСТ Р 51177-98 — Оборудование для воздушных линий. Общие технические условия
ГОСТ Р 52209-2004 — Рукава в сборе для газовых горелок и газовых приборов. Общие технические требования и методы испытаний
ГОСТ Р 52630-2006 — Сосуды и аппараты стальные сварные.Общие технические условия
ГОСТ Р 52630-2012 — Сосуды и аппараты стальные сварные. Основные Характеристики.
ГОСТ Р 53279-2009 — Оборудование пожарное. Огнестойкие соединительные головки. Общие технические требования. Методы тестирования
ГОСТ Р 53325-2009 — Техника пожарная. Средства пожарной автоматики. Общие технические требования. Методы испытаний
ГОСТ Р 55472-2013 — Системы газораспределения. Требования к газораспределительным сетям. Часть 0. Общие
ГОСТ Р 55509-2013 — Арматура трубопроводная.Металлы, используемые в производстве трубопроводной арматуры. Основные требования к выбору материалов
Основные требования к выбору материалов
ОДМ 218.7.002-2008: Методические указания по определению глубины проникновения иглы для вязкого нефтяного дорожного битума
ОДМ 218.7.004-2008: Рекомендации по методике определения температуры размягчения вязкого нефтяного дорожного битума на кольце и шаре
ПБ 03-384-00 — Правила проектирования, изготовления и приемки сосудов и аппаратов стальных сварных
ПБ 03-576-03 — Правила устройства и безопасной эксплуатации сосудов под давлением
.ПБ 10-115-96 — Правила устройства и безопасной эксплуатации сосудов под давлением
.ПНСТ 3-2012 — Дороги автомобильные общего пользования.Битумы нефтяные дорожные вязкие. Определение проникновения иглы
ПНСТ 4-2012 — Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Определение точки размягчения. Метод кольца и шарика
РД 24.207.01-90 — Арматура трубопроводная. Зависимость среднего срока службы гидрозатвора от удельного давления на уплотнительную поверхность
РД 26-07-274-89 — Арматура трубопроводная. Кольца латунные для задвижек из серого чугуна по Ру до 1.0 МПа (10 кгс
Кольца латунные для задвижек из серого чугуна по Ру до 1.0 МПа (10 кгс
СП 42-101-2003 — Система газораспределения общего положения и строительства из стальных и полиэтиленовых труб
СТ ЦКБА 002-2003 — Арматура трубопроводная.Защелки. Методика расчета силы
СТ ЦКБА 005.1-2003 — Арматура трубопроводная. Металлы, используемые в арматурной промышленности. Часть 1. Основные требования к выбору материалов
СТ ЦКБА 005.3-2004 — Арматура трубопроводная. Металлы, используемые в арматурной промышленности. Часть 3. Зарубежные материалы и их отечественные аналоги
СТ ЦКБА 005.3-2009 — Арматура трубопроводная. Металлы, используемые в арматурной промышленности. Часть 3. Зарубежные материалы и их отечественные аналоги
СТ ЦКБА 051-2008 — Арматура трубопроводная.Отливки из цветных сплавов. Технические требования
СТ ЦКБА 060-2008 — Арматура трубопроводная. Ходовые резьбовые пары. Основные размеры, технические требования
СТ ЦКБА 068-2008 — Арматура трубопроводная. Запорная арматура с металлическим уплотнением. Технические требования
Запорная арматура с металлическим уплотнением. Технические требования
ГОСТ 10944-2019 — Краны ручные и запорные для систем водяного отопления зданий. Общие технические условия
ГОСТ 11614-2019 — Краны смывные полуавтоматические. Технические характеристики
ГОСТ 12393-2019 — Арматура контактная рельсовая линейная.Общие технические условия
ПНСТ 375-2019 — Баллоны газовые. Технические требования и испытания автоматических газовых запорных клапанов для баллонов со сжиженным газом
Клиентов, которые просматривали этот товар, также просматривали:
|
 Общие технические условия
Общие технические условия Общие технические условия
Общие технические условия Метод испытания на ударную вязкость при низкой, комнатной и высокой температуре
Метод испытания на ударную вязкость при низкой, комнатной и высокой температуреВАШ ЗАКАЗ ПРОСТО!
Белорусское законодательство. com — ведущая в отрасли компания со строгими стандартами контроля качества, и наша приверженность точности, надежности и аккуратности является одной из причин, почему некоторые из крупнейших мировых компаний доверяют нам обеспечение своей национальной нормативно-правовой базы и перевод критических, сложных и конфиденциальная информация.
com — ведущая в отрасли компания со строгими стандартами контроля качества, и наша приверженность точности, надежности и аккуратности является одной из причин, почему некоторые из крупнейших мировых компаний доверяют нам обеспечение своей национальной нормативно-правовой базы и перевод критических, сложных и конфиденциальная информация.
Наша нишевая специализация — локализация национальных нормативных баз данных, включающих: технические нормы, стандарты и правила; государственные законы, кодексы и постановления; а также кодексы, требования и инструкции агентств РФ.
У нас есть база данных, содержащая более 220 000 нормативных документов на английском и других языках для следующих 12 стран: Армения, Азербайджан, Беларусь, Казахстан, Кыргызстан, Молдова, Монголия, Россия, Таджикистан, Туркменистан, Украина и Узбекистан.
Размещение заказа
Выберите выбранный вами документ, перейдите на «страницу оформления заказа» и выберите желаемую форму оплаты. Мы принимаем все основные кредитные карты и банковские переводы.Мы также принимаем PayPal и Google Checkout для вашего удобства. Свяжитесь с нами для любых дополнительных договоренностей (договорные соглашения, заказ на поставку и т. Д.).
Мы принимаем все основные кредитные карты и банковские переводы.Мы также принимаем PayPal и Google Checkout для вашего удобства. Свяжитесь с нами для любых дополнительных договоренностей (договорные соглашения, заказ на поставку и т. Д.).
После размещения заказа он будет проверен и обработан в течение нескольких часов, но в редких случаях — максимум 24 часа.
Документ / веб-ссылка для товаров на складе будет отправлена вам по электронной почте, чтобы вы могли загрузить и сохранить ее для своих записей.
Если товары отсутствуют на складе (поставка сторонних поставщиков), вы будете уведомлены о том, для каких товаров потребуется дополнительное время.Обычно мы поставляем такие товары менее чем за три дня.
Как только заказ будет размещен, вы получите квитанцию / счет, который можно будет заполнить для отчетности и бухгалтерского учета. Эту квитанцию можно легко сохранить и распечатать для ваших записей.
Гарантия лучшего качества и подлинности вашего заказа
Ваш заказ предоставляется в электронном формате (обычно это Adobe Acrobat или MS Word).
Мы всегда гарантируем лучшее качество всей нашей продукции.Если по какой-либо причине вы не удовлетворены, мы можем провести совершенно БЕСПЛАТНУЮ ревизию и редактирование приобретенных вами продуктов. Кроме того, мы предоставляем БЕСПЛАТНЫЕ обновления нормативных требований, если, например, документ имеет более новую версию на дату покупки.
Гарантируем подлинность. Каждый документ на английском языке сверяется с оригинальной и официальной версией. Мы используем только официальные нормативные источники, чтобы убедиться, что у вас самая последняя версия документа, причем все из надежных официальных источников.
Законы Армении | Официальная нормативная библиотека — ГОСТ 1440-78
Продукт содержится в следующих классификаторах:
ПромЭксперт »
РАЗДЕЛ I. ТЕХНИЧЕСКИЙ РЕГЛАМЕНТ »
V Тестирование и контроль »
3 Деятельность испытательных лабораторий »
3.2 Техническое оснащение испытательных лабораторий »
3. 2.2 Измерительные приборы и испытательное оборудование »
3.2.2.7 Приборы для определения состава, состояния и свойств веществ »
2.2 Измерительные приборы и испытательное оборудование »
3.2.2.7 Приборы для определения состава, состояния и свойств веществ »
Классификатор ISO » 17 МЕТРОЛОГИЯ И ИЗМЕРЕНИЯ. ФИЗИЧЕСКИЕ ЯВЛЕНИЯ » 17.060 Измерения объема, массы, плотности, вязкости »
Национальные стандарты » 17 МЕТРОЛОГИЯ И ИЗМЕРЕНИЯ. ФИЗИЧЕСКИЕ ЯВЛЕНИЯ » 17.060 Измерения объема, массы, плотности, вязкости »
Национальные стандарты для сомов » Последнее издание » P Измерительные приборы.Средства автоматизации и вычислительной техники » П6 Приборы для определения состава, состояния и свойств веществ » П66 Аппараты, посуда и лабораторное оборудование из стекла, фарфора и кварца »
В качестве замены:
ГОСТ 1440-42 — Прибор для определения проникновения нефтепродуктов
.
Ссылка на документ:
ГОСТ 11501-78 — Битум нефтяной. Метод определения глубины проникновения иглы пенетрометра
ГОСТ 15836-79 — Мастика битумно-резиновая изоляционная.Технические характеристики
ГОСТ 21179-2000 — Воск пчелиный. Технические характеристики
ГОСТ 21179-90 — Воск пчелиный. Технические характеристики
ГОСТ 2488-79 — Церезин. Технические характеристики
ГОСТ 25374-82 — Воск пчелиный экстракционный. Технические характеристики
ГОСТ 25593-83 — Пасты алмазные
.ГОСТ 25771-83 — Парафин и церезины. Метод пенетрации иглой
ГОСТ 25945-87 — Материалы и изделия полимерные строительные незатвердевающие. Методы испытаний
ГОСТ 25945-98 — Материалы и изделия уплотнительные строительные полимерные незатвердевающие.Методы испытаний
ГОСТ 33136-2014 — Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Метод определения глубины проникновения иглы пенетрометра
ГОСТ 5346-78 — Смазки пластичные. Методы определения пенетрации коническим пенетрометром
Методы определения пенетрации коническим пенетрометром
ГОСТ 7142-74 — Смазки консистентные. Методы определения сепарации нефти
ОДМ 218.7.002-2008: Методические указания по определению глубины проникновения иглы для вязкого нефтяного дорожного битума
ОСТ 6-15-1525-86 — Пластилин детский.Технические условия
ПНСТ 3-2012 — Дороги автомобильные общего пользования. Битумы нефтяные дорожные вязкие. Определение проникновения иглы
СТБ 1087-97 — Пластилин детский. Технические характеристики
ВСН 27-76 — Технические инструкции по применению битумных шламов для устройства защитных слоев автомобильных дорог
Клиентов, которые просматривали этот товар, также просматривали:
|
 Оценки
Оценки Определение плотности жидкостей и твердых тел
Определение плотности жидкостей и твердых тел Методы испытаний
Методы испытаний Общие требования безопасности
Общие требования безопасностиВАШ ЗАКАЗ ПРОСТО!
ArmeniaLaws.com — ведущая в отрасли компания со строгими стандартами контроля качества, и наша приверженность точности, надежности и аккуратности является одной из причин, почему некоторые из крупнейших мировых компаний доверяют нам обеспечение своей национальной нормативно-правовой базы и перевод критических, сложных и конфиденциальная информация.
Наша нишевая специализация — локализация национальных нормативных баз данных, включающих: технические нормы, стандарты и правила; государственные законы, кодексы и постановления; а также кодексы, требования и инструкции агентств РФ.
У нас есть база данных, содержащая более 220 000 нормативных документов на английском и других языках для следующих 12 стран: Армения, Азербайджан, Беларусь, Казахстан, Кыргызстан, Молдова, Монголия, Россия, Таджикистан, Туркменистан, Украина и Узбекистан.
Размещение заказа
Выберите выбранный вами документ, перейдите на «страницу оформления заказа» и выберите желаемую форму оплаты. Мы принимаем все основные кредитные карты и банковские переводы.Мы также принимаем PayPal и Google Checkout для вашего удобства. Свяжитесь с нами для любых дополнительных договоренностей (договорные соглашения, заказ на поставку и т. Д.).
После размещения заказа он будет проверен и обработан в течение нескольких часов, но в редких случаях — максимум 24 часа.
Документ / веб-ссылка для товаров на складе будет отправлена вам по электронной почте, чтобы вы могли загрузить и сохранить ее для своих записей.
Если товары отсутствуют на складе (поставка сторонних поставщиков), вы будете уведомлены о том, для каких товаров потребуется дополнительное время.Обычно мы поставляем такие товары менее чем за три дня.
Как только заказ будет размещен, вы получите квитанцию / счет, который можно будет заполнить для отчетности и бухгалтерского учета. Эту квитанцию можно легко сохранить и распечатать для ваших записей.
Гарантия лучшего качества и подлинности вашего заказа
Ваш заказ предоставляется в электронном формате (обычно это Adobe Acrobat или MS Word).
Мы всегда гарантируем лучшее качество всей нашей продукции.Если по какой-либо причине вы не удовлетворены, мы можем провести совершенно БЕСПЛАТНУЮ ревизию и редактирование приобретенных вами продуктов. Кроме того, мы предоставляем БЕСПЛАТНЫЕ обновления нормативных требований, если, например, документ имеет более новую версию на дату покупки.
Гарантируем подлинность. Каждый документ на английском языке сверяется с оригинальной и официальной версией. Мы используем только официальные нормативные источники, чтобы убедиться, что у вас самая последняя версия документа, причем все из надежных официальных источников.
| № | Параметры | Типичный для | Метод испытаний | |||
| Альфабит 40 Мост | Альфабит 40 Аэро | Альфабит 60 Аэро | Альфабит 90 Аэро | |||
| 1 | Глубина проникновения иглы, 0.  1 мм, мин., 1 мм, мин., при температуре: | ГОСТ 33136 | ||||
| при 25 ℃ | 40 | 40 | 60 | 90 | ||
| при 0 ℃ | 25 | 25 | 32 | 40 | ||
| 2 | Предел прочности, см, не менее при температуре: | ГОСТ 33138 | ||||
| при 25 ℃ | 30 | 30 | 35 | 40 | ||
| при 0 ℃ | 10 | 10 | 15 | 20 | ||
| 3 | Температура размягчения на кольце и шаре, ℃, мин. | 70 | 70 | 65 | 65 | ГОСТ 33142 |
| 4 | Температура хрупкости, ℃, не более .  | -18 | -18 | -22 | -27 | ГОСТ 33143 |
| 5 | Эластичность,%, не менее при температуре: | ГОСТ Р 52056 п.6.2 | ||||
| при 25 ℃ | 85 | 85 | 90 | 90 | ||
| при 0 ℃ | 70 | 70 | 70 | 75 | ||
| 6 | Температура вспышки, ℃, мин. | 235 | 235 | 235 | 235 | ГОСТ 33141 |
| 7 | Тяга с мрамором или песком | Обеспечено контрольным образцом 1 | ГОСТ 11508, метод А | |||
| 8 | Однородность | Однородный | ГОСТ Р 52056 п.  6.1 6.1 | |||
| 9 | Динамическая вязкость при 135 ℃, Па * с | Для набора статистических данных | ГОСТ 33137 с добавлением п.7.1 настоящего стандарта | |||
| 10 | Устойчивость к старению | ГОСТ 33140 | ||||
| Изменение температуры размягчения после старения, ℃, макс. (по абсолютной величине) | 5 | 5 | 5 | 5 | ГОСТ 33142 | |
| Температура хрупкости после старения, ℃, не выше | -15 | -15 | -20 | -25 | ГОСТ 33136 | |
| Эластичность при 25 ℃, %, мин. | 80 | 80 | 85 | 85 | ГОСТ Р 52056 п.  6.2 6.2 | |
| 11 | Устойчивость при хранении, через 72 ч | ГОСТ ЕN 13399 | ||||
| Изменение температуры размягчения на , ℃ | ≤5 | ≤5 | ≤5 | ≤5 | ГОСТ Р 33142 | |
Резюме
Введение Polycomb репрессивный комплекс 1 (PRC1) представляет собой модульно собранный мультисубъединичный комплекс, который контролирует программную экспрессию генов посредством воздействия на эпигенетические метки гистонов.PRC1 определяется наличием ферментативного ядра RING-PCGF, основная активность которого заключается в моноубиквитинировании h3A на лизине 119 (Blackledge et al., 2020; Gao et al., 2012; Tamburri et al., 2020). PRC1 может собираться в виде 6 различных комплексов (PRC1.1-PRC1.6), каждый из которых определяется конкретным линкерным белком PCGF, который связывает ферменты RING1 / RNF2 с отдельным набором вспомогательных субъединиц, которые определяют его механизм нацеливания на хроматин и регуляторную функцию гена ( Gao et al. , 2012; Scelfo et al., 2019). Из этих шести подкомплексов PRC1 только PRC1.1, как известно, соматически мутирует при раке (Sashida et al., 2019).
, 2012; Scelfo et al., 2019). Из этих шести подкомплексов PRC1 только PRC1.1, как известно, соматически мутирует при раке (Sashida et al., 2019).
В PRC1.1 ферментное ядро связано с помощью PCGF1 с KDM2B, BCOR, BCORL1 и вспомогательными субъединицами SKP1 и USP7 (Gao et al., 2012). BCOR и BCORL1 периодически мутируют при гематологических злокачественных новообразованиях, включая острые и хронические лейкозы. При остром миелоидном лейкозе (AML) мутации BCOR и BCORL1 были зарегистрированы у 5-10% пациентов и связаны с предшествующим диагнозом миелодиспластического синдрома (MDS) и плохими клиническими исходами после химиотерапии (Branford et al., 2018; Damm et al., 2013; Гроссманн и др., 2011; Линдсли и др., 2015, 2017; Папаэммануил и др., 2016; Terada et al., 2018). При хроническом миелоидном лейкозе (ХМЛ) мутации BCOR связаны с запущенным заболеванием и клинической резистентностью к ингибиторам, направленным на BCR-ABL (Branford et al., 2018). Связанные с лейкемией соматические мутации BCOR и BCORL1 приводят к преждевременным стоп-кодонам, но механизмы, с помощью которых эти мутации изменяют сборку и функцию комплекса PRC1. 1 и вносят вклад в лейкемогенез, неизвестны.
1 и вносят вклад в лейкемогенез, неизвестны.
Основным препятствием для нашего понимания влияния мутаций BCOR и BCORL1 на биологию PRC1.1 является отсутствие информации относительно сборки субъединиц in vivo, локализации хроматина и эффектов на регуляцию целевого гена. Исследования in vitro показали, что взаимодействие BCOR / BCORL1 с PCGF1 необходимо для последующей сборки с KDM2B (Wong et al., 2016, 2020), но последствия потери BCOR / BCORL1 на целостность комплекса in vivo не были показаны. Исследования дополнительно приписывают различные функции другим структурным доменам BCOR, таким как BCL6-связывающий и AF9-связывающий домены (Ghetu et al., 2008; Schmidt et al., 2020; Srinivasan et al., 2003), повышая вероятность того, что соматические мутации могут иметь гетерогенные эффекты на активность PRC1.1 или могут опосредовать эффекты на неканонические функции PRC1.1, которые не зависят от активности убиквитинирования h3A. Наконец, было показано, что PRC1.1 широко рекрутируется на неметилированные промоторы островков CpG через домен KDM2B CXXC (Farcas et al. , 2012; He et al., 2013; Wu et al., 2013), но подмножество связанных локусы, которые являются истинными функциональными мишенями PRC1.1, не были определены при лейкемии.
, 2012; He et al., 2013; Wu et al., 2013), но подмножество связанных локусы, которые являются истинными функциональными мишенями PRC1.1, не были определены при лейкемии.
Здесь мы комбинируем геномный, биохимический и функциональный подходы для анализа роли мутаций BCOR и BCORL1 в миелоидных злокачественных новообразованиях. Мы определяем влияние мутаций отдельных субъединиц на сборку комплекса, нацеливание на хроматин и экспрессию целевого гена. Эти исследования предоставляют ключевую информацию о нормальной функции PRC1.1 и последствиях мутаций, связанных с заболеванием человека, тем самым раскрывая прямую механистическую связь между эпигенетическим репрограммированием и активацией сигнальных путей, что приводит к фенотипу лекарственной устойчивости.
Результаты Мутации BCOR и BCORL1 вызывают потерю белка или усечения C-концевого белка и в значительной степени одновременно возникают при миелоидных злокачественных новообразованиях. Для определения частоты и спектра мутаций PRC1. 1 при AML мы выполнили целевое секвенирование генов, кодирующих все девять субъединиц PRC1.1. в образцах костного мозга 433 больных ОМЛ. Наиболее часто мутировали BCOR (26 из 433, 6%) и BCORL1 (8 из 433, 1,8%) (рис. 1A). Мы идентифицировали одну мутацию в определяющей комплекс субъединице PCGF1 (0,2%), одну мутацию в KDM2B (0.2%) и отсутствие мутаций в генах, кодирующих другие PRC1.1-специфические дополнительные субъединицы (SKP1, USP7) или ферментативное ядро пан-PRC1 (RYBP, RING1, RNF2) (рис. 1A, таблица S1). Эти генетические находки показывают, что избирательное изменение PRC1.1 с помощью мутаций BCOR и BCORL1, но не изменение PRC1 более глобально, вносит вклад в патогенез миелоидных злокачественных новообразований.
1 при AML мы выполнили целевое секвенирование генов, кодирующих все девять субъединиц PRC1.1. в образцах костного мозга 433 больных ОМЛ. Наиболее часто мутировали BCOR (26 из 433, 6%) и BCORL1 (8 из 433, 1,8%) (рис. 1A). Мы идентифицировали одну мутацию в определяющей комплекс субъединице PCGF1 (0,2%), одну мутацию в KDM2B (0.2%) и отсутствие мутаций в генах, кодирующих другие PRC1.1-специфические дополнительные субъединицы (SKP1, USP7) или ферментативное ядро пан-PRC1 (RYBP, RING1, RNF2) (рис. 1A, таблица S1). Эти генетические находки показывают, что избирательное изменение PRC1.1 с помощью мутаций BCOR и BCORL1, но не изменение PRC1 более глобально, вносит вклад в патогенез миелоидных злокачественных новообразований.
Список вариантов генов, кодирующих PRC1.1 субъединицы.
 Мутации
Мутации(A) Частота генных мутаций в отдельных субъединицах PRC1.1 в когорте из 433 пациентов с AML. Число над столбиками указывает количество пациентов с мутациями. См. Также Таблицу S1.
(B) Столбиковая диаграмма, представляющая распределение заболеваний мутаций BCOR и / или BCORL1 в когорте из 3955 последовательных пациентов с диагнозом миелоидных злокачественных новообразований и 1774 человек с гематологическими аномалиями, но без гематологического диагноза злокачественных новообразований.
(C) График вулкана, представляющий ассоциации мутаций генов BCOR и BCORL1 у пациентов с AML (верхняя панель) или MDS (нижняя панель). Ось x показывает величину ассоциации (отношение шансов log2), а ось y — значение q -log10. Каждый кружок представляет собой обозначенный мутантный ген. Гены в верхнем правом квадранте значительно совпадали.
Гены в верхнем правом квадранте значительно совпадали.
(D) Графики рыб, представляющие предполагаемую клональную динамику на основе секвенирования. См. Также Таблицу S2.
(E) График Lollipop, представляющий мутации сдвига рамки считывания BCOR или BCORL1.Мутации, чувствительные к NMD (черные кружки; NMD +) и нечувствительные к NMD (красные кружки; NMD-), были определены с помощью инструмента прогнозирования NMDetective-B (Lindeboom et al., 2019). PTC с баллами NMD <0,25 считались нечувствительными к NMD, PTC с баллами NMD> 0,25 считались чувствительными к NMD. Высококонсервативные области BCOR и BCORL1 определяли с помощью Blastp (инструмент поиска базового локального выравнивания белок-белок) (Altschul et al., 1997). NMD: нонсенс-опосредованный распад мРНК. PTC: кодон преждевременной терминации.
(F) Вестерн-блоттинг (верхняя панель) лизатов целых клеток из образцов пациентов с AML с мутацией BCOR WT или BCOR. Антитело BCOR RRID: AB_2716801 использовали для определения уровней белка BCOR (эпитоп: аминокислоты 1035-1230, (Gearhart et al., 2006)). Pt.3 содержит чувствительную к NMD мутацию BCOR (NMD +; p.S1439fs * 44, прогнозируемый размер: 161 кДа). Pt.5 содержит мутацию BCOR, нечувствительную к NMD (NMD-; p.L1647fs * 3, прогнозируемый размер: 180 кДа). Уровни экспрессии гена BCOR (нижняя панель), представленные в виде нормализованного количества DESeq2 в BCOR WT (Pt.1) или BCOR PUFD-Tr (Pt.5) первичные образцы пациентов с ОМЛ (среднее ± стандартное отклонение, n = 2 технических дубликата для каждого состояния).
Антитело BCOR RRID: AB_2716801 использовали для определения уровней белка BCOR (эпитоп: аминокислоты 1035-1230, (Gearhart et al., 2006)). Pt.3 содержит чувствительную к NMD мутацию BCOR (NMD +; p.S1439fs * 44, прогнозируемый размер: 161 кДа). Pt.5 содержит мутацию BCOR, нечувствительную к NMD (NMD-; p.L1647fs * 3, прогнозируемый размер: 180 кДа). Уровни экспрессии гена BCOR (нижняя панель), представленные в виде нормализованного количества DESeq2 в BCOR WT (Pt.1) или BCOR PUFD-Tr (Pt.5) первичные образцы пациентов с ОМЛ (среднее ± стандартное отклонение, n = 2 технических дубликата для каждого состояния).
Частоты вариативных аллелей (VAF) вариантов, обнаруженные у 4 пациентов мужского пола в двух временных точках. nd: не определено.
В отдельной когорте из 3955 последовательных пациентов с диагнозом миелоидные злокачественные новообразования в одном учреждении мы обнаружили, что мутации BCOR и BCORL1 были наиболее распространены при AML (121/1551, 7.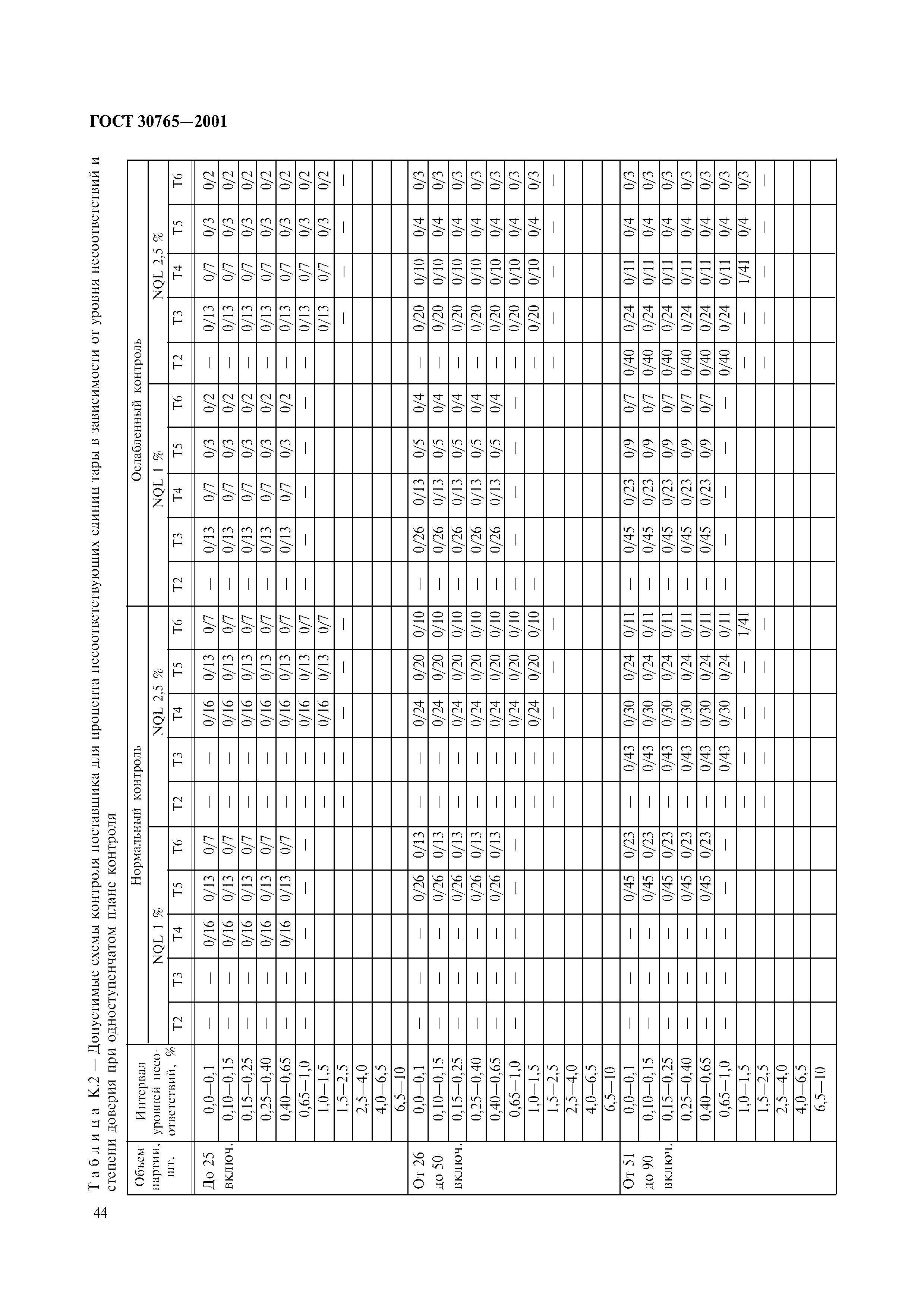 8%) и MDS (36/942, 5,0%), за которыми следуют MDS / MPN (10/343, 2,9%), Ph-MPN (8/868, 0,9%) и хроническая фаза CML (2/251, 0,8%). %) (Рисунок 1B). Мутации BCOR или BCORL1 не наблюдались у 1774 последовательных пациентов с гематологическими аномалиями, но не обнаружено гематологических злокачественных новообразований. Мутации BCOR и BCORL1 в значительной степени совпадают как при МДС (LR = 4,19, q <0,001), так и при AML (LR = 3,57, q <0,0001), что указывает на то, что они могут иметь кооперативные эффекты в патогенезе заболевания (рис. 1C). В соответствии с этой гипотезой, 45.7% (16/35) пациентов с ОМЛ с мутацией BCORL1 также имели мутацию BCOR, а серийные образцы от пациентов с одновременными мутациями BCOR и BCORL1 показали, что мутация BCORL1 возникла субклонально из уже существовавшей мутации BCOR (рис. 1D, таблица S2). .
8%) и MDS (36/942, 5,0%), за которыми следуют MDS / MPN (10/343, 2,9%), Ph-MPN (8/868, 0,9%) и хроническая фаза CML (2/251, 0,8%). %) (Рисунок 1B). Мутации BCOR или BCORL1 не наблюдались у 1774 последовательных пациентов с гематологическими аномалиями, но не обнаружено гематологических злокачественных новообразований. Мутации BCOR и BCORL1 в значительной степени совпадают как при МДС (LR = 4,19, q <0,001), так и при AML (LR = 3,57, q <0,0001), что указывает на то, что они могут иметь кооперативные эффекты в патогенезе заболевания (рис. 1C). В соответствии с этой гипотезой, 45.7% (16/35) пациентов с ОМЛ с мутацией BCORL1 также имели мутацию BCOR, а серийные образцы от пациентов с одновременными мутациями BCOR и BCORL1 показали, что мутация BCORL1 возникла субклонально из уже существовавшей мутации BCOR (рис. 1D, таблица S2). .
Соматические мутации BCOR (n = 256) и BCORL1 (n = 90) в этой когорте были распределены по экзонам с 4 по 15, и большинство (311 из 346, 89,9%) привело к введению кодонов преждевременной терминации (Таблица S3). Среди всех мутаций сдвига рамки считывания BCOR и BCORL1 90.Было предсказано, что 8% (423/466) вызовут снижение экспрессии белка через нонсенс-опосредованный распад мРНК (NMD-чувствительные мутации, NMD +), а 9,2% (43/466), как ожидается, вызовут стабильную экспрессию усеченных на С-конце белков, в которых отсутствует часть связывающего домена PCGF1 PUFD (мутации, нечувствительные к NMD, NMD-). Распределение мутаций BCOR и BCORL1 в двух текущих когортах, а также в наших ранее опубликованных когортах MDS (Lindsley et al., 2017) и AML (Lindsley et al., 2015) показано на рисунке 1E.В соответствии с прогнозами, мы обнаружили экспрессию усеченного белка BCOR PUFD в первичных бластах AML, несущих нечувствительную к NMD мутацию (Pt.5 BCOR p.L1647fs * 3), тогда как уровень белка BCOR был заметно снижен в первичных бластах AML, несущих чувствительные к NMD мутации (Pt.3 BCOR p.S1439fs * 44) (Рисунок 1F). Связанные с заболеванием мутации BCOR, таким образом, приводят либо к потере экспрессии белка, либо к экспрессии усеченного на С-конце белка, у которого отсутствует интактный домен PUFD.
Среди всех мутаций сдвига рамки считывания BCOR и BCORL1 90.Было предсказано, что 8% (423/466) вызовут снижение экспрессии белка через нонсенс-опосредованный распад мРНК (NMD-чувствительные мутации, NMD +), а 9,2% (43/466), как ожидается, вызовут стабильную экспрессию усеченных на С-конце белков, в которых отсутствует часть связывающего домена PCGF1 PUFD (мутации, нечувствительные к NMD, NMD-). Распределение мутаций BCOR и BCORL1 в двух текущих когортах, а также в наших ранее опубликованных когортах MDS (Lindsley et al., 2017) и AML (Lindsley et al., 2015) показано на рисунке 1E.В соответствии с прогнозами, мы обнаружили экспрессию усеченного белка BCOR PUFD в первичных бластах AML, несущих нечувствительную к NMD мутацию (Pt.5 BCOR p.L1647fs * 3), тогда как уровень белка BCOR был заметно снижен в первичных бластах AML, несущих чувствительные к NMD мутации (Pt.3 BCOR p.S1439fs * 44) (Рисунок 1F). Связанные с заболеванием мутации BCOR, таким образом, приводят либо к потере экспрессии белка, либо к экспрессии усеченного на С-конце белка, у которого отсутствует интактный домен PUFD.
Список вариантов BCOR (n = 256) и BCORL1 (n = 90).
Чтобы определить влияние мутаций BCOR и BCORL1 на сборку и функцию комплекса PRC1.1, мы создали изогенные клеточные линии K562 со сдвигом рамки считывания BCOR и / или Мутации BCORL1, которые напоминали спектр мутаций, которые мы наблюдали у пациентов с миелоидными злокачественными новообразованиями (рис. 2А).Предсказанные NMD-чувствительные мутации BCOR в экзоне 7, экзоне 9 или экзоне 10 приводили к потере экспрессии белка BCOR (BCOR KO ), тогда как предсказанные нечувствительные к NMD мутации BCOR в экзоне 14 вызывали экспрессию усеченных белков с С-конца (BCOR PUFD -Tr ) (Рисунки 2B, S1A и S1B). Мутации BCORL1 в экзоне 8 или экзоне 9 привели к потере BCORL1 (BCORL1 KO ) (Фигуры 2C и S1C).
(A) Обзор изогенных клеточных линий BCOR и BCORL1 (K562). Леденцы представляют собой одноклеточные клоны с отдельными мутациями BCOR (верхняя панель) или мутациями BCORL1 (нижняя панель). Мутации, чувствительные к NMD (черные кружки; NMD +) и нечувствительные к NMD (красные кружки; NMD-), были определены с помощью инструмента прогнозирования NMDetective-B (Lindeboom et al., 2019). PTC с баллами NMD <0,25 считались нечувствительными к NMD, PTC с баллами NMD> 0,25 считались чувствительными к NMD.A / L: домены ANK / L. PD: домены PUFD. См. Также рисунки S1A и S1C.
(B) BCOR- и BCORL1-Co-IP-вестерн-блот анализ в клетках BCOR-WT (WT), BCOR PUFD-Tr (Tr) или BCOR KO (KO) K562 (ядерные экстракты, эндогенные белки) ). Короткая (90 кДа) и длинная (152 кДа) изоформы KDM2B отмечены стрелками (звездочка указывает неспецифическую полосу).
(C) Экспрессия гена PCGF1 (нормализованное количество DESeq2) в WT, BCORL1 KO , BCOR PUFD-Tr , BCOR KO , BCOR PUFD-Tr / BCORL1 KO16 K или BCORL1 KO16 KO или BCORL1 KO16 KO Клеточные линии BCORL1 KO K562 представлены как среднее ± стандартное отклонение (верхняя панель).Нижняя панель показывает соответствующий вестерн-блот-анализ ядерных экстрактов.
(D) Вестерн-блоттинг (лизаты цельных клеток) субъединиц PRC1 в клетках пациентов с AML BCOR WT (Pt.6) или BCOR KO (Pt.7, BCOR p.R1164 *).
(E) Co-IP-вестерн-блот анализ экзогенно экспрессируемых BCOR HA-PUFD (1635-1755aa; левая панель) или BCOR HA-CT (1462-1755aa; правая панель) в BCOR KO / BCORL1 KO К562 клетки. Клетки BCOR KO / BCORL1 KO , трансдуцированные контрольным вектором, использовали в качестве отрицательного контроля.
(F) Экспрессия гена BCOR или BCORL1 (нормализованное количество DESeq2) в клетках WT, BCORL1 KO или BCOR KO K562, представленных как среднее ± стандартное отклонение.
(G) Относительные уровни экспрессии генов BCOR и BCORL1 в образцах первичного AML или клеточных линиях AML, как указано. Значения FPKM образцов AML (beatAML n = 507, TARGET n = 532, LAML n = 156) были получены из исследовательской сети TCGA: https://www.cancer.gov/tcga. Значения RPKM клеточной линии AML были загружены из Карты зависимости рака Института Брод (Broad Institute Cancer Dependency Map, 2018).
(H) Медианная экспрессия гена (нормализованное количество DESeq2) BCOR и BCORL1 в гемопоэтических стволовых / клетках-предшественниках и зрелых клонах, как указано. Подсчеты РНК-секвенированных линий клеток, отсортированных по FACS, были получены от Corces et al. (Corces et al., 2016).
(A) Вестерн-блоттинг-анализ лизатов цельных клеток из WT, BCOR KO или BCOR PUFD-Tr K562 клонов единичных клеток (относящихся к фигуре 2A).Антитела, распознающие N-концевые (аминокислоты 1-300, RRID: AB_2721913) или C-концевые (аминокислоты 1396-1755, RRID: AB_22
(B) Экспрессия гена BCOR (нормализованное количество DESeq2) в клетках WT, BCOR KO или BCOR PUFD-Tr K562, представленная как среднее ± стандартное отклонение.
(C) Вестерн-блоттинг лизатов целых клеток от WT или BCORL1 KO K562 клонов одиночных клеток (относящихся к Фигуре 2A).Прогнозируемая молекулярная масса BCORL1-WT: 1785 а.о. / 190 кДа.
(D) PCGF1- и KDM2B-Co-IP-вестерн-блот-анализ (ядерные экстракты) в клетках WT, BCOR KO (KO) или BCOR PUFD-Tr (Tr) K562. Короткая (90 кДа) и длинная (152 кДа) изоформы KDM2B отмечены стрелками (звездочка указывает неспецифическую полосу).
(E) BCOR-Co-IP-вестерн-блот-анализ в клетках WT или BCOR PUFD-Tr (Tr) MOLM14. Клетки BCOR PUFD-Tr MOLM14 представляют собой основную популяцию с частотами отступов экзона 14 BCOR> 90%, определенными секвенированием по Сэнгеру и анализом TIDE (Brinkman et al., 2014). Короткая (90 кДа) и длинная (152 кДа) изоформы KDM2B отмечены стрелками (звездочка указывает неспецифическую полосу).
(F) Вестерн-блот-анализ ядерных экстрактов из WT, BCORL1 KO , BCOR PUFD-Tr , BCOR KO , BCOR PUFD-Tr / BCORL1 KO или BCOR / BCORL 916 KO K562 клеточных линий (относится к фиг. 2C; общий контроль загрузки: LMNB1). Короткая (90 кДа) и длинная (152 кДа) изоформы KDM2B отмечены стрелками (звездочка указывает неспецифическую полосу).
(G) Уровни экспрессии гена PCGF1 (нормализованные значения DESeq2) в BCOR-WT (Pt.1) или BCOR PUFD-Tr (Tr, Pt.5; BCOR p.L1647fs * 3) в клетках пациентов с AML представлены как среднее ± SD (верхняя панель). Нижняя панель показывает соответствующий вестерн-блот-анализ лизатов целых клеток.
(H) KDM2B-Co-IP-вестерн-блот-анализ в клетках BCOR KO / BCORL1 KO K562, экспрессирующих HA-BCOR-CT (1462-1755aa). Клетки BCOR KO / BCORL1 KO , трансдуцированные контрольным вектором, использовали в качестве отрицательного контроля.
(I) Вестерн-блоттинг связанных и несвязанных IP-фракций анализа BCOR- или BCORL1-Co-IP в клетках WT или BCOR KO K562. IgG использовали в качестве отрицательного контроля.
Структурное моделирование показало, что гетеродимеры BCOR-PCGF1 и BCORL1-PCGF1 образуются отдельно (Junco et al., 2013; Wong et al., 2016), но неизвестно, рекрутируются ли эти гетеродимеры одновременно в отдельные PRC1.1. комплексов или независимо от различных комплексов in vivo. Чтобы различать эти возможности, мы выполнили реципрокный BCOR- и BCORL1-IP и вестерн-блоттинг в клетках WT (рис. 2B).Мы обнаружили, что BCOR и BCORL1 отсутствовали в IP реципрокного паралога, указывая на то, что BCOR-PRC1.1 и BCORL1-PRC1.1 являются взаимоисключающими комплексами. Кроме того, взаимодействия BCORL1 с PCGF1 и KDM2B были стабильными в отсутствие BCOR, что позволяет предположить, что комплексы BCOR-PRC1.1 и BCORL1-PRC1.1 собираются независимо.
Ранее in vitro было показано, что BCORL1-PUFD опосредует прямое взаимодействие с PCGF1, которое требуется для последующего взаимодействия с KDM2B (Wong et al., 2016, 2020). Напротив, недавнее исследование чЭСК показало, что BCOR-PUFD необходим для связывания PCGF1, но не KDM2B (Wang et al., 2018). Используя BCOR-Co-IP, мы обнаружили, что усечения BCOR-PUFD нарушают взаимодействия с PCGF1, не влияя на связывание BCOR с KDM2B (рис. 2B). Мы подтвердили наши результаты с помощью взаимных PCGF1- и KDM2B-Co-IP (рисунок S1D). В соответствии с нашими выводами в клетках K562, усечения BCOR-PUFD в клетках MOLM14 нарушали связывание с PCGF1, но не с KDM2B (рисунок S1E). Эти данные указывают на то, что BCOR-PUFD выполняет важную функцию в связывании PCGF1, который избирательно нарушается даже у пациентов с высоко экспрессируемыми C-концевыми мутациями, усекающими BCOR.
BCOR и BCORL1 контролируют уровни белка PCGF1 посредством прямых стабилизирующих взаимодействий.PCGF1-RAWUL, как сообщается, требует прямого взаимодействия с BCOR / BCORL1-PUFD для стабильности in vitro (Junco et al., 2013), но влияние этого взаимодействия на in vivo Сборка комплекса PRC1.1 или стабильность полноразмерного белка PCGF1 не были продемонстрированы. Поэтому мы проанализировали влияние мутаций BCOR и BCORL1 на содержание PCGF1 в клетках. В условиях BCOR KO / BCORL1 KO мы наблюдали полную потерю белка PCGF1, несмотря на отсутствие значительных различий в уровне мРНК PCGF1 (рис. 2C).Напротив, одновременная инактивация BCOR / BCORL1 не влияла на количество других PRC1.1-специфичных субъединиц (KDM2B, USP7, SKP1), субъединиц ферментативного ядра пан-PRC1 (RING1, RNF2, RYBP, CBX8), белков PCGF (BMI1). , PCGF3, PCGF5) или PRC2 (EZh3) (рисунок S1F). Уровень белка PCGF1 был снижен в образцах первичного AML с мутациями BCOR (рисунки 2D и S1G).
Мы наблюдали подобный избирательный эффект на уровень белка PCGF1 в клетках BCOR PUFD-Tr / BCORL1 KO (рис. 2C), что указывает на то, что белок BCOR, лишенный PUFD, не может стабилизировать PCGF1.Чтобы проверить, достаточно ли C-концевых PCGF1-связывающих доменов BCOR для восстановления взаимодействий BCOR-PCGF1 и стабильности белка PCGF1 in vivo, мы экспрессировали HA-меченные домены BCOR-PUFD или BCOR-CT в BCOR KO / BCORL1 КО ячеек (рис. 2Е). Экспрессии BCOR-PUFD было достаточно для образования стабильного гетеродимера с PCGF1 и восстановления взаимодействия с RNF2 (рис. 2E). В соответствии с предыдущими выводами (Wang et al., 2018; Wong et al., 2020), домен BCOR-CT, содержащий домены ANK / линкер и PUFD, аналогичным образом восстанавливает стабильное взаимодействие с PCGF1, но также связан с KDM2B (Рисунки 2E и S1H).Таким образом, домен PUFD был необходим и достаточен для связывания и стабилизации белка PCGF1, тогда как ANK / линкер был необходим для совместного рекрутирования KDM2B.
Единственное нарушение BCOR (BCOR KO или BCOR PUFD-Tr ) вызвало только частичное снижение уровней белка PCGF1, а единственное нарушение BCORL1 (BCORL1 KO ) не повлияло на изобилие белка PCGF1 (Рисунок 2C), указывает на то, что постоянная экспрессия паралогичных BCORL1 или BCOR компенсирует потерю BCOR или BCORL1, соответственно.Мы предположили, что различное влияние BCOR по сравнению с потерей BCORL1 на общий уровень белка PCGF1 было связано с различиями в уровне экспрессии паралога на исходном уровне. В соответствии с этой гипотезой, BCOR экспрессировался более высоко, чем BCORL1 в клетках WT K562 (рис. 2F). В более широком смысле, BCOR был преимущественно экспрессируемым паралогом в других клеточных линиях BCOR / BCORL1 WT AML (KG1, OCIAML3, MV4-11 и MOLM13), в первичных образцах AML и во всем нормальном гематопоэзе, включая стволовые клетки и клетки-предшественники, и зрелые клетки крови всех клонов (Рисунки 2G и 2H).В соответствии с более высокими уровнями экспрессии BCOR, BCOR вносил вклад в большинство взаимодействий PCGF1 в анализе Co-IP (рисунок S1I). Мы также не наблюдали компенсаторного увеличения экспрессии гена BCORL1 или уровней белка в мутантных клетках BCOR (рис. 2F). Вместе наши данные показывают, что распространенность PCGF1 напрямую связана с составным уровнем уровней экспрессии BCOR и BCORL1 и что снижение уровней PCGF1 в отдельных BCOR или мутантных клетках BCORL1 пропорционально базовым уровням экспрессии оставшихся паралогов.
Потеря взаимодействия BCOR-PCGF1 или BCORL1-PCGF1 нарушает сборку комплекса за счет отделения ферментативного ядра от KDM2BМы показали, что общий эффект инактивации BCOR и BCORL1 заключается в дестабилизации уровней белка PCGF1. Следовательно, мы затем попытались определить влияние прямой потери PCGF1 на сборку комплекса PRC1.1. Для создания изогенных клеточных линий PCGF1 KO мы ввели мутации в домен PCGF1-RING или в область, которая ранее была идентифицирована как важный контактный сайт для взаимодействий PCGF1-PUFD (рисунок 3A) (Wong et al., 2016). Потеря PCGF1 не повлияла на стабильность белка BCOR или BCORL1 (рис. 3A).
biorxiv; 2021.03.08.433705v1 / FIG3F3fig3 Рисунок 3. Потеря(A) Вестерн-блоттинг ядерных экстрактов клеток WT или PCGF1 KO K562. Мутации PCGF1 показаны на схеме ниже. KO1: PCGF1-p.I60fs *. KO2: PCGF1-p. (I60fs *; p.Y163fs *). KO3: PCGF1-p.Y163del. Антитело PCGF1 RRID: AB_2721914 использовали для определения уровней белка PCGF1 (эпитоп: аминокислоты 128–247).
(B) Обогащение субъединиц PRC1.1 в масс-спектрометрическом анализе BCOR-IP в клетках WT или PCGF1 KO K562, представленных как изменение в log2 раза по сравнению с контролем BCOR KO . n = 2 (процесс повторяется для каждого условия). См. Также рисунки S2A-C.
(C) KDM2B-Co-IP-вестерн-блот-анализ (фракции элюирования, ядерные экстракты) в клетках WT или BCORL1 KO K562 (левая панель) и в WT, BCOR PUFD-Tr (Tr) или BCOR КО (КО) Ячейки К562 (правая панель).Короткая (90 кДа) и длинная (152 кДа) изоформы KDM2B отмечены стрелками (звездочка указывает неспецифическую полосу).
(D) KDM2B-Co-IP-вестерн-блот-анализ (ядерные экстракты) в WT, BCOR PUFD-Tr / BCORL1 KO (Tr / KO) или BCOR KO / BCORL1 KO (DKO) К562 клетки. Короткая (90 кДа) и длинная (152 кДа) изоформы KDM2B отмечены стрелками (звездочка указывает неспецифическую полосу).
(A) Обогащение партнеров по связыванию BCOR в анализе BCOR-IP MS. Графики вулканов показывают -log10 (p-значение) против log2-кратного изменения. n = 2 (процесс повторяется для каждого условия). Субъединицы, определяющие PRC1.1 (BCOR, KDM2B, PCGF1), выделены красным, ферментное ядро (RING1, RNF2, RYBP, CBX8) — синим, а вспомогательные субъединицы (SKP1, USP7) — черным.
(B) Обогащение субъединиц PRC1.1 в анализе BCOR-IP MS в клетках WT, PCGF1 KO или BCOR KO K562, представленных как изменение в log2 раза по сравнению с контролем IgG.n = 2 (процесс повторяется для каждого условия).
(C) BCOR-Co-IP-вестерн-блоттинг (ядерные экстракты) в клетках WT или PCGF1 KO K562. Антитело PCGF1 RRID: AB_2721914 использовали для определения уровней белка PCGF1 (эпитоп: аминокислоты 128–247). Короткая (90 кДа) и длинная (152 кДа) изоформы KDM2B отмечены стрелками (звездочка указывает неспецифическую полосу).
(D) Фракции элюирования BCOR-Co-IP-вестерн-блоттинга в первичных клетках пациента AML BCOR WT (верхняя панель) или клеточных линиях OCI-AML3 и KG1 (нижняя панель).Короткая (90 кДа) и длинная (152 кДа) изоформы KDM2B отмечены стрелками.
(E) Фракции элюирования BCORL1-Co-IP-вестерн-блоттинга в клетках WT K562. Черная линия показывает результаты двух независимых экспериментов.
(F) KDM2B-Co-IP-вестерн-блот-анализ в клетках MOLM14 WT или BCOR PUFD-Tr (Tr). Клетки BCOR PUFD-Tr MOLM14 представляют собой основную популяцию с частотами отступов экзона 14 BCOR> 90%, определенными секвенированием по Сэнгеру и анализом TIDE (Brinkman et al., 2014). Короткая (90 кДа) и длинная (152 кДа) изоформы KDM2B отмечены стрелками (звездочка указывает неспецифическую полосу).
(G) BCOR- и KDM2B-Co-IP-вестерн-блот-анализ в WT, BCOR KO / BCORL1 KO (DKO) или BCOR KO / BCORL1 KO клеток, дополненных WT-BCOR (DKO BCOR ). Экзогенный полноразмерный WT-BCOR (изоформа BCOR 1) стабильно экспрессировался в клетках DKO BCOR с помощью лентивирусной трансдукции.
Чтобы идентифицировать BCOR-взаимодействующие белки в присутствии или в отсутствие PCGF1, мы иммунопреципитировали BCOR и проанализировали комплекс с помощью масс-спектрометрии (MS) в клетках WT или PCGF1 KO .В клетках WT основными партнерами по взаимодействию BCOR были PRC1.1-определяющие и вспомогательные субъединицы (PCGF1, KDM2B, USP7 и SKP1) и субъединицы ферментного ядра пан-PRC1 (RING1 / RNF2, RYBP и CBX8) (рис. 3B, S2A и S2B). Мы подтвердили эти результаты на клетках K562 с помощью IP-вестерн-блоттинга (рисунок S2C) и подтвердили взаимодействия BCOR с основными субъединицами PRC1.1 в первичных образцах пациентов с AML и в клеточных линиях OCI-AML3 и KG1 (рисунок S2D). Взаимодействия BCORL1 с PRC1.1-специфичными и пан-PRC1 субъединицами отражали взаимодействия BCOR (рисунок S2E).В клетках PCGF1 KO мы наблюдали потерю связывания BCOR с ферментативным ядром, включая RING1, RNF2, RYBP и CBX8 (Фигуры 3B и S2A-S2C).
Однако отсутствие PCGF1 не нарушает взаимодействия BCOR с KDM2B, SKP1 и USP7, указывая на то, что PCGF1 важен для связывания ферментативного ядра с комплексом, но не для опосредования взаимодействий BCOR-KDM2B.
Определить, оказывает ли непрямая потеря белка PCGF1 посредством мутаций BCOR и BCORL1 такой же эффект на PRC1.1 как прямая потеря PCGF1 посредством делеции гена, мы выполнили KDM2B-Co-IP в BCORL1 KO , BCOR PUFD-Tr , BCOR KO , BCOR PUFD-Tr / BCORL1 KO и BCOR КО / BCORL1 КО ячеек. По отдельности ни BCOR, ни BCORL1 мутации не вызывали полного нарушения взаимодействий KDM2B с субъединицами PRC1.1 (Figure 3C), что согласуется с нашим наблюдением, что BCOR-PRC1.1 и BCORL1-PRC1.1 образуют независимые комплексы с функцией избыточного адаптера в PRC1.1 сборка. Мы наблюдали аналогичные результаты в клетках BCOR PUFD-Tr MOLM14 (фигура S2F). Одновременное разрушение BCOR и BCORL1 привело к полному разъединению ферментативного ядра PRC1 от KDM2B, что отражается в потере связывания RING1, RNF2, RYBP и CBX8 (рис. 3D). Это открытие было идентичным в клетках, лишенных экспрессии белка BCOR (BCOR KO / BCORL1 KO ) и клетках, экспрессирующих усеченный PUFD BCOR (BCOR PUFD-Tr / BCORL1 KO ), что дополнительно подтверждает специфичность и абсолютное требование. для BCOR-PUFD в PRC1.1 сложная сборка. Чтобы подтвердить этот вывод, мы дополнили клетки BCOR KO / BCORL1 KO посредством экзогенной экспрессии полноразмерного BCOR и продемонстрировали восстановление уровней белка PCGF1 и взаимодействия KDM2B с ферментативными субъединицами ядра (рисунок S2G).
KDM2B необходим для рекрутирования BCOR-PRC1.1 и BCORL1-PRC1.1 на неметилированные островковые промоторы CpGДля определения локализации комплексов BCOR- и BCORL1-PRC1.1 на хроматине мы выполнили BCOR, BCORL1, KDM2B и RNF2 ChIP-seq в ячейках K562.BCOR и BCORL1 связывались преимущественно с промоторными областями (рисунки 4A и S3A), и распределения связывания BCOR и BCORL1 в сайтах начала транскрипции (TSS) были значительно коррелированы (R 2 = 0,7375, p <0,0001; Рисунок 4B). Связывание BCORL1 не изменилось в клетках BCOR KO по сравнению с клетками WT (фиг. 4A и 4B), а гены, связанные с BCOR и BCORL1, были совместно заняты субъединицами PRC1.1 KDM2B и RNF2 (фиг. 4A). Вместе эти результаты показывают, что BCOR-PRC1.1 и BCORL1-PRC1.1 комплексы независимо рекрутируются в общий набор целевых локусов.
biorxiv; 2021.03.08.433705v1 / FIGS3F10figs3 Рисунок S3.(A) Общее количество пиков BCOR и BCORL1 (порог: -log10 (q-значение)> 10) в клетках WT K562 (верхняя панель). Распределение пиков BCOR и BCORL1 в клетках WT K562 в соответствии с их геномным положением (нижняя панель).
(B) Диаграммы рассеяния, показывающие взаимосвязь между сигналами BCOR-ChIP-seq при TSS +/- 5kb в BCOR PUFD-Tr vs.Клетки WT K562 (верхняя панель) и клетки PCGF1 KO по сравнению с клетками WT K562 (нижняя панель).
Коэффициенты корреляции Пирсона и значения p (двусторонние), как указано.
(C) Общее количество пиков BCOR (порог: -log10 (q-value)> 10) в BCOR KO , BCOR PUFD-Tr , PCGF1 KO или KDM2B KO K562 ячеек (верхний панель). Распределение пиков BCOR в соответствии с их геномным положением (нижняя панель).
(D) Тепловая карта представляет собой обогащение BCOR-ChIP BCOR WT (Pt.1) или BCOR PUFD-Tr (Pt.5; BCOR p.L1647fs * 3) первичные образцы пациентов с AML при TSS +/- 5kb.
(E) BCOR-Co-IP-вестерн-блот-анализ (ядерные экстракты) в клетках WT или KDM2B KO K562. Короткая (90 кДа) и длинная (152 кДа) изоформы KDM2B отмечены стрелками (звездочка указывает неспецифическую полосу).
(F) Средние сигналы BCOR ChIP-seq (левая панель) и сигналы BCORL1 ChIP-seq (правая панель) в клетках WT K562 при промоторах, отличных от CGI (n = 7414), или промоторах CGI, разделенных на квантили на основе количества CpG на CGI (Q1-4 n = 2200; Q5 n = 2199).CGI: остров CpG.
(G) Профили средней плотности сигналов BCOR-ChIP в первичных образцах пациентов с AML BCOR WT (Pt.1) при богатых CpG (n = 10 851) или бедных CpG (n = 7352) промоторах (левая панель). Средние сигналы BCOR ChIP-seq в первичных образцах пациентов с AML BCOR WT на промоторах, не относящихся к CGI (n = 7414), или промоторах CGI, разделенные на квантили на основе количества CpG на CGI (Q1-4 n = 2200; Q5 n = 2199) .
(H) Средние сигналы BCOR ChIP-seq в клеточных линиях KG1, OCI-AML3 или MV4-11 AML на промоторах, отличных от CGI (n = 7414), или промоторах CGI, разделенных на квантили на основе количества CpG на CGI (Q1- 4 n = 2200; Q5 n = 2199).
(I) Средние сигналы BCOR- и BCORL1-ChIP-seq на промоторах CGI, подразделенные на основе средних процентов метилирования на CGI.
(A) Тепловые карты представляют собой обогащение BCOR-, BCORL1-, KDM2B- и RNF2-ChIP на сайтах начала транскрипции (TSS) +/- 5kb в клетках WT, BCORL1 KO или BCOR KO K562. Показатели сигнала BCOR-ChIP в клетках WT K562 использовали для предварительного определения PRC1.1 связанный (n = 11934) и несвязанный PRC1.1 (n = 6639) промоторы. Все тепловые карты сортируются и ранжируются на основе обогащения BCOR-ChIP в клетках WT.
(B) Диаграммы рассеяния, показывающие взаимосвязь между сигналами BCORL1- и BCOR-ChIP-seq при TSS +/- 5kb в клетках WT K562 (верхняя панель). Диаграммы рассеяния, показывающие взаимосвязь между сигналами BCORL1-ChIP-seq в клетках BCOR KO и WT K562 (нижняя панель). Биологические реплики условий BCOR KO нанесены черным (Rep1) или синим (Rep2) цветом.Коэффициенты корреляции Пирсона и значения p (двусторонние), как указано.
(C) Тепловые карты обогащения BCOR-ChIP-seq на PRC1.1 связанном (n = 11934) и несвязанном PRC1.1 (n = 6639) промоторах гена в BCOR KO , BCOR PUFD-Tr , PCGF1 КО , или КДМ2Б КО ячеек. Все тепловые карты сортируются и ранжируются на основе сигналов BCOR-ChIP в клетках WT (рис. 4A).
(D) Профили средней плотности сигналов BCOR-ChIP и BCORL1-ChIP в клетках WT K562 на промоторах с высоким содержанием CpG (n = 10,851) или с промоторами с низким содержанием CpG (n = 7352).См. Также рисунок S3F.
(E) Отслеживание данных геномного покрытия для BCOR-ChIP-seq и BCORL1-ChIP-seq на неметилированных (FGFR1) или метилированных (TLX2) промоторах островков CpG. Процент метилирования CpG-сайтов указан цветовым градиентом.
(F) Схема набора PRC1.1 в условиях BCOR PUFD-Tr , PCGF1 KO и KDM2B KO . BCOR стабильно рекрутируется для генов-мишеней в условиях BCOR PUFD-Tr и PCGF1 KO , но не в клетках KDM2B KO , что указывает на то, что для PRC1 требуется KDM2B, но не ферментное ядро.1 набор лейкозных клеток.
В человеческих ESC связывание PRC1.1 с большинством генов-мишеней, как сообщается, зависит от RING1 / RNF2 (Wang et al., 2018), тогда как в мышиных ESC рекрутирование PRC1.1 в значительной степени опосредуется KDM2B (Farcas et al. al., 2012; He et al., 2013; Wu et al., 2013). Это указывает на то, что механизм рекрутирования PRC1.1 может зависеть от типа клеток. Чтобы определить дифференциальную потребность в ферментативном ядре PRC1 и KDM2B в привлечении комплекса PRC1.1 в лейкозных клетках человека, мы выполнили BCOR-ChIP-seq в клетках WT или клетках, дефицитных по разным PRC1.1 субъединицы. Как в клетках BCOR PUFD-Tr , так и в клетках PCGF1 KO , где BCOR селективно отделен от ферментативного ядра, сохраняя при этом связывание с KDM2B, количество пиков BCOR было аналогичным по сравнению с WT (рисунки 4C, S3A и S3C) и Распределение связывания BCOR в TSS значимо коррелировало с обогащением BCOR в WT (WT против BCOR PUFD-Tr , R 2 = 0,8949, p <0,0001 и WT против PCGF1 KO , R 2 = 0,9413 , p <0,0001; рисунок S3B).Мы подтвердили этот вывод на первичных образцах пациентов, подтвердив, что усеченный BCOR поддерживает связывание с генами-мишенями (рисунок S3D).
Чтобы проверить необходимость KDM2B в нацеливании PRC1.1, мы создали изогенные клеточные линии KDM2B KO путем введения мутаций сдвига рамки считывания выше ДНК-связывающего домена CXXC, что привело к потере как длинной, так и короткой изоформ. В этих условиях KDM2B KO BCOR сохранял нормальные взаимодействия с PRC1.1-специфическими субъединицами (USP7 и PCGF1) и ферментным ядром PRC1, но терял KDM2B-зависимое связывание с SKP1 (рисунок S3E).В отсутствие KDM2B мы наблюдали глобальную потерю связывания хроматина BCOR, которая была эквивалентна условиям отрицательного контроля BCOR KO (Фигуры 4C, 4E и S3C), что указывает на то, что нацеливание PRC1.1 на хроматин полностью зависит от KDM2B. Сообщалось, что KDM2B связывается с ДНК посредством своего домена цинковых пальцев CXXC, который распознает неметилированные CpG-островки (CGI) (Farcas et al., 2012; He et al., 2013; Wu et al., 2013). В соответствии с механизмом KDM2B-зависимого рекрутирования мы обнаружили, что связывание BCOR и BCORL1 было обогащено промоторами CGI и положительно коррелировало с содержанием CpG на промоторах CGI в клетках K562 (рисунки 4D и S3F), а также в образцах первичных пациентов с ОМЛ и в клеточных линиях лейкемии KG1, OCI-AML3 и MV411 (Рисунки S3G и S3H).Используя расширенное бисульфитное секвенирование с пониженной репрезентативностью (ERRBS) в клетках WT K562, мы обнаружили, что занятость BCOR и BCORL1 отрицательно коррелировала с уровнями метилирования ДНК в CGI (Фигуры 4E и S3I). Вместе эти результаты показывают, что KDM2B, но не PCGF1 или ферментное ядро, важен для рекрутирования BCOR- и BCORL1-PRC1.1 на неметилированные CpG-островки при лейкемии (рис. 4F).
Рекрутирование ферментативного ядра PRC1.1 необходимо для репрессии целевого генаДалее мы исследовали состояние хроматина PRC1.1-связанные и PRC1.1-несвязанные гены с использованием h4K27ac, h4K4me3, h4K27me3 и h3AK119ub ChIP-seq и ATAC-seq. Среди генов, связанных с PRC1.1, 78,2% (9336 из 11934) имели активное состояние хроматина, отраженное высокими уровнями h4K27ac и h4K4me3, высокой доступностью хроматина и высокой базовой экспрессией генов (Рисунки 5A и S4A). 21,8% (2598 из 11934) генов, связанных с PRC1.1, имели репрессированное состояние хроматина с высоким уровнем h4K27me3, высоким уровнем h3AK119ub и низкой экспрессией гена или ее отсутствием (Фигуры 5A и S4A). Напротив, PRC1.1-несвязанные гены (n = 6269) в основном имели неактивное состояние хроматина, связанное с уплотненным хроматином и отсутствием экспрессии генов (Фигуры 5A и S4A). Мы наблюдали аналогичное распределение связывания BCOR в клеточных линиях KG1, OCI-AML3 и MV4-11 и в первичных клетках пациентов с AML (Фигуры S4A-S4D).
biorxiv; 2021.03.08.433705v1 / FIGS4F11figs4 Рисунок S4.(A) Уровни экспрессии генов (нормализованное количество DESeq2) активных, репрессированных или неактивных генов в клеточных линиях K562, KG1, OCI-AML3 или MV411.Оценки сигналов BCOR- и h4K27ac-ChIP в клетках K562 WT использовали для определения активного, репрессированного и неактивного состояний хроматина (как показано на фиг. 5A и S4C). Линия в середине прямоугольника представляет собой медианное значение, края прямоугольника представляют 25-й и 75-й процентили, а усы показывают 5-й и 95-й процентили.
(B) Уровни экспрессии генов (нормализованное количество DESeq2) активных, репрессированных или неактивных генов в первичных клетках AML BCOR WT (Pt.1). Оценки сигналов BCOR- и h4K27ac-ChIP использовали для определения активного, подавленного и неактивного состояний хроматина (как показано на рисунке S4D).Линия в середине прямоугольника представляет собой медианное значение, края прямоугольника представляют 25-й и 75-й процентили, а усы показывают 5-й и 95-й процентили.
(C) Тепловые карты и профили сигналов ChIP-seq BCOR, модификаций гистонов и сигналов доступности хроматина (ATAC) в клетках KG-1, OCI-AML3 и MV4-11 при активных, репрессированных или неактивных промоторах гена ( TSS +/- 5кб).
(D) Тепловые карты и профили сигналов ChIP-seq BCOR, модификаций гистонов и сигналов доступности хроматина (ATAC) в первичных клетках AML BCOR WT (Pt.1) на активных, репрессированных или неактивных промоторах гена (TSS +/- 5kb).
(E) Уровни экспрессии генов (log2-трансформированные нормализованные значения DESeq2) активных (n = 9335), репрессированных (n = 2597) или неактивных (n = 6268) генов (как определено на рисунке 5A) в WT или PRC1.1 мутантные клетки K562. Линия в середине прямоугольника представляет собой медианное значение, края прямоугольника представляют 25-й и 75-й процентили, а усы показывают 5-й и 95-й процентили. Для статистического анализа использовался критерий суммы рангов Вилкоксона. n.s. р> 0,05, * р <0.05, ** p <0,01, *** p <0,001, **** p <0,0001.
(F) Диаграммы Венна представляют собой перекрытия активированных генов в мутантных клетках PRC1.1 по сравнению с контролем дикого типа (padj <0,05, кратное изменение> 2). В этот анализ были включены только гены, которые на исходном уровне находились в подавленном состоянии хроматина.
(G) Графики в виде прямоугольников и усов, представляющие уровни экспрессии (log2-трансформированные нормализованные значения DESseq2) генов, не отвечающих на PRC1.1 (n = 1799) и реагирующих на PRC1.1 (n = 632), у мутанта K562 WT или PRC1.1 ячеек (относящихся к Фигуре 5D).Линия в середине прямоугольника представляет собой медианное значение, края прямоугольника представляют 25-й и 75-й процентили, а усы показывают 5-й и 95-й процентили. Для статистического анализа использовался критерий суммы рангов Вилкоксона. n.s. p> 0,05, * p <0,05, ** p <0,01, *** p <0,001, **** p <0,0001.
(H) Профили средней плотности гистоновых сигналов ChIP-seq в клетках WT K562 в генах, отвечающих на PRC1.1 (n = 632) или нечувствительных к PRC1.1 (n = 1799).
(I) Тепловые карты, представляющие сигналы h3AK119ub- и SUZ12-ChIP при TSS +/- 5kb в ячейках WT или BCOR KO / BCORL1 KO .
(A) Тепловые карты и графики профиля обогащения ChIP-seq BCOR, модификаций гистонов и сигналов доступности хроматина (ATAC), как указано. Показатели сигнала BCOR-ChIP в клетках K562 WT использовали для предварительного определения промоторов, связанных с PRC1.1 (n = 11934). Показатели сигнала BCOR- и h4K27ac-ChIP в клетках K562 WT использовали для определения активных, репрессированных и неактивных промоторов генов.Данные K562 h4K4me3-ChIP-seq были загружены из архива считывания последовательностей (SRA; SRR341173). Остальные наборы данных были созданы в этом исследовании. См. Также рисунок S4A.
(B) Число дифференциально экспрессируемых генов в мутанте PRC1.1 по сравнению с клетками WT K562 (кратное изменение> 2, padj <0,05). Дифференциально экспрессируемые гены были подразделены на основе их состояний хроматина, определенных на фиг. 5A.
(C) Столбчатая диаграмма представляет процентное соотношение генов с повышенной регуляцией в мутантных клетках PRC1.1 по отношению к общему количеству генов в каждом состоянии хроматина.
(D) Неконтролируемый иерархический кластерный анализ уровней экспрессии PRC1.1 связанных репрессированных генов в WT (n = 7) или PRC1.1 мутантных клетках K562 (BCORL1 KO (n = 3), BCOR PUFD-Tr (n = 2), BCOR KO (n = 7), BCOR KO / BCORL1 KO (n = 3), BCOR PUFD-Tr / BCORL1 KO (n = 2), KDM2B КО (n = 4), PCGF1 KO (n = 2)). Гены сгруппированы в гены, отвечающие на PRC1.1 (n = 632) или не отвечающие на PRC1.1 (n = 1799).Гены, реагирующие на PRC1.1, сгруппированы в гены C1 (n = 65) или C2 (n = 567). Уровни экспрессии генов показаны в виде относительных преобразованных значений VST. См. Также рисунок S4G.
(E) Профили средней плотности сигналов BCOR-, BCORL1- и RNF2-ChIP в клетках WT K562 в генах, отвечающих на PRC1.1 (n = 632) или нечувствительных к PRC1.1 (n = 1799).
(F) Число подсчетов CpG на PRC1.1-ответных (n = 632) или нечувствительных промоторах CGI (n = 1799).
**** p <0,0001, непарный t-критерий. Линия в середине прямоугольника представляет собой медианное значение, края прямоугольника представляют 25-й и 75-й процентили, а усы показывают 5-й и 95-й процентили.
(G) Графики в виде прямоугольников и усов, представляющие гистоновые ChIP- и ATAC-seq сигналы в генах, отвечающих на PRC1.1 (n = 632), нормализованные по сигналам в генах, не отвечающих на PRC1.1 (n = 1799) в WT или BCOR КО / BCORL1 КО ячеек. Линия в середине прямоугольника представляет собой медианное значение, края прямоугольника представляют 25-й и 75-й процентили, а усы показывают 2,5-й и 97,5-й процентили.
(H) Профили средней плотности сигналов SUZ12-ChIP в клетках WT или BCOR KO / BCORL1 KO K562 на PRC1.1 реагирующий (n = 632) или нечувствительный к PRC1.1 ген (n = 1799).
(I) Данные о покрытии ChIP-seq и ATAC-seq отслеживают PRC1.1-чувствительный ген JAG1 в клетках WT или BCOR KO / BCORL1 KO K562.
Связывание с хроматином PRC1 само по себе не коррелирует напрямую с регуляторной активностью (Farcas et al., 2012; Klose et al., 2013). Таким образом, мы определили набор функциональных мишеней PRC1.1, анализируя влияние полной инактивации PRC1.1 на глобальную экспрессию генов по сравнению с PRC1.1 клетки WT. Мы обнаружили, что преобладающим последствием была активация гена, независимо от механизма комплексной инактивации, включая полную потерю PRC1.1 (BCOR KO / BCORL1 KO : 85,6%, 1481/1731), нарушение рекрутирования PRC1.1 в хроматина (KDM2B KO : 83,3%, 1380/1656) и избирательное разобщение ферментативного ядра PRC1 (BCOR PUFD-Tr / BCORL1 KO : 81,8%, 1060/1296 и PCGF1 KO : 92,3% , 756/819) (Рисунок 5B). Доля дифференциально экспрессируемых генов была самой высокой среди генов с репрессированным состоянием хроматина на исходном уровне, а гены с активным и неактивным состоянием хроматина демонстрировали относительно небольшие изменения (Рисунки 5B и S4E).Действительно, среди набора репрессированных PRC1.1-связанных генов мы наблюдали значительную активацию гена в BCOR KO / BCORL1 KO (29,8%), KDM2B KO (26,2%), BCOR PUFD-Tr / BCORL1 KO (21,5%) и PCGF1 KO (18,9%) (фиг. 5C), и имело место высокое перекрытие репрессированных генов с повышенной регуляцией во всех мутантных условиях PRC1.1 (фиг. S4F). Вместе эти результаты указывают на то, что нарушение PRC1.1 вызывает селективную дерепрессию генов-мишеней.
Чтобы идентифицировать набор генов-мишеней PRC1.1 среди генов с репрессированным хроматином на исходном уровне, мы выполнили неконтролируемую иерархическую кластеризацию экспрессии генов в WT и всех мутантных клетках PRC1.1. Мы определили два основных кластера генов: гены, отвечающие на PRC1.1 (n = 632) и не отвечающие на PRC1.1 гены (n = 1799) (рис. 5D). Уровни экспрессии генов, отвечающих за PRC1.1, были сильно увеличены во всех неактивных условиях PRC1.1 (BCOR PUFD-Tr / BCORL1 KO , BCOR KO / BCORL1 KO , PCGF1 KO , KDM2B KO , KDM2B ) по сравнению с контролем дикого типа, тогда как PRC1.1 нечувствительные гены показали сходные уровни экспрессии в мутантных клетках WT и PRC1.1 (рисунок S4G). Чтобы идентифицировать параметры, которые были связаны с реактивностью PRC1.1, мы оценили занятость PRC1.1 и модификации гистонов в генах, отвечающих или нечувствительных к PRC1.1. Гены, отвечающие на PRC1.1, показали более высокие уровни h3AK119ub и h4K27me3, чем гены, не отвечающие на PRC1.1 (рисунок S4H). Мы также наблюдали более сильное связывание BCOR, BCORL1 и RNF2 у реагирующих генов по сравнению с нечувствительными генами, которые коррелировали с более высокими уровнями CpG на промоторах CGI (Фигуры 5E и 5F), указывая на то, что PRC1.1 был необходим для поддержания репрессии определенного набора генов-мишеней, богатых CpG.
Недавние исследования показали, что отложение h3AK119ub с помощью неканонических PRC1 важно для рекрутирования PRC2 и поддержания репрессии целевых генов (Blackledge et al., 2020; Fursova et al., 2019; Tamburri et al., 2020). Мы предположили, что потеря PRC1.1 нарушает отложение h3AK119ub, что приводит к эпигенетической активации PRC1.1-чувствительных генов. Чтобы проверить эту гипотезу, мы выполнили h3AK119ub, h4K27me3, h4K27ac и ATAC-seq в клетках BCOR KO / BCORL1 KO .Мы наблюдали снижение уровней h3AK119ub и h4K27me3 в генах, отвечающих за PRC1.1, в клетках BCOR KO / BCORL1 KO по сравнению с клетками WT, в то время как уровни h4K27ac и доступность хроматина были увеличены (рисунки 5G и 5I). Нарушение PRC1.1 и снижение отложения h3AK119ub в генах, отвечающих за PRC1.1, были дополнительно связаны со снижением занятости PRC2 (Рисунки 5H и 5I), тогда как глобальные уровни h3AK119ub или PRC2 не были нарушены (Рисунок S4I). Эти данные предполагают, что ферментативная активность PRC1.1 поддерживает активную репрессию специфических генов-мишеней за счет отложения h3AK119ub и стабильного рекрутирования PRC2.
PRC1.1 регулирует клеточные сигнальные программы при лейкемии человекаЧтобы идентифицировать PRC1.1-зависимые программы транскрипции в лейкозных клетках, мы выполнили анализ обогащения набора генов дифференциально экспрессируемых генов в клетках BCOR KO / BCORL1 KO по сравнению с BCOR WT / BCORL1 WT клеток. Дифференциально экспрессируемые гены в условиях BCOR KO / BCORL1 KO были обогащены сигнальными путями, регулирующими плюрипотентность стволовых клеток (KEGG hsa04550), включая передачу сигналов FGF / FGFR, передачу сигналов PI3K-Akt и пути передачи сигналов TGF-ß, 6 (Фигуры 6B, SA5A) (Фигуры 6B, SA5A). , и S5B), что соответствует роли PRC1.1 в регуляции сигнальных программ стволовых клеток. Мы также наблюдали обогащение сигнальных путей, вовлеченных в рак (KEGG hsa05200), таких как сигнальные пути RAS и MAPK (Фигуры 6A, S5A и S5B).
biorxiv; 2021.03.08.433705v1 / FIGS5F12figs5 Рисунок S5.(A) Избыточно представлены пути передачи сигналов KEGG на основании анализа обогащения набора генов дифференциально экспрессируемых генов в BCOR KO / BCORL1 KO по сравнению с клетками WT K562.NES: нормализованная оценка обогащения.
(B) Анализ обогащения пути KEGG генов, чувствительных к PRC1.1 (n = 632), с использованием gProfiler (g: GOSt) (Raudvere et al., 2019).
(C) Перепредставленные сигнальные пути KEGG на основе анализа обогащения набора генов дифференциально экспрессируемых генов в BCOR mut (n = 22) по сравнению с BCOR WT (n = 216) первичными образцами AML из исследования beatAML (Tyner et al. др., 2018). NES: нормализованная оценка обогащения.
(D) График вулкана, представляющий изменения экспрессии генов с усиленной регуляцией в BCOR mut (n = 22) по сравнению с первичными образцами AML BCOR WT (n = 216) из исследования beatAML (Tyner et al., 2018). Гены, представленные в сигнальных путях KEGG, регулирующих плюрипотентность стволовых клеток (hsa04550) или пути KEGG при раке (hsa05200), выделены красным. Изменения кратности Log2 1 и padj 0,05 показаны пунктирными линиями. 275 генов были значительно усилены более чем в 2 раза в клетках BCOR mut по сравнению с клетками BCOR WT .
(E) Анализ обогащения набора генов KEGG дифференциально экспрессируемых генов в BCOR PUFD-Tr по сравнению с образцами первичного AML BCOR WT .Верхний график представляет величину log2-кратных изменений для каждого гена. Нижний график показывает оценку обогащения (ES) (левая панель). Резюме чрезмерно представленных сигнальных путей KEGG на основе анализа обогащения набора генов дифференциально экспрессируемых генов в BCOR PUFD-Tr по сравнению с образцами первичного AML BCOR WT (правая панель). NES: нормализованная оценка обогащения.
(A) Анализ обогащения набора генов KEGG (GSEA) дифференциально экспрессируемых генов в BCOR KO / BCORL1 KO по сравнению с клетками WT K562. Верхний график представляет величину log2-кратных изменений для каждого гена. Нижний график показывает оценку обогащения (ES). См. Также рисунок S5A.
(B) График вулкана, представляющий изменения экспрессии генов активированных генов в BCOR KO / BCORL1 KO по сравнению с клетками WT K562. Гены, представленные в сигнальных путях KEGG, регулирующих плюрипотентность стволовых клеток (hsa04550) или пути KEGG при раке (hsa05200), выделены красным.Изменения кратности Log2 1 и padj 0,05 показаны пунктирными линиями. В клетках BCOR KO / BCORL1 KO была значительно повышена более чем в 2 раза регуляция генов 1642 по сравнению с контролем WT.
(C) Анализ обогащения набора генов KEGG дифференциально экспрессируемых генов в BCOR mut (n = 22) по сравнению с BCOR WT (n = 216) образцов пациентов с AML из исследования beatAML (Tyner et al., 2018) . Верхний график представляет величину log2-кратных изменений для каждого гена.Нижний график показывает оценку обогащения (ES). См. Также рисунок S5C.
(D) Графики в виде прямоугольников и усов, представляющие уровни экспрессии (log2-преобразованные нормализованные значения DESeq2) в первичных образцах AML BCOR WT (n = 216) или BCOR mut (n = 22) (Tyner et al., 2018) . Линия в середине прямоугольника представляет собой медианное значение, края прямоугольника представляют 25-й и 75-й процентили, а усы показывают 5-й и 95-й процентили.
(E) Дорожки данных покрытия ChIP-seq PRC1.1 гены-мишени JAG1 и FOXO1 в клетках BCOR WT (Pt.1) или BCOR PUFD-Tr (Pt.5) (левая панель). Соответствующие уровни экспрессии генов JAG1 и FOXO1 (нормализованные значения DESeq2, n = 2, технические повторы; правая панель).
Чтобы подтвердить наши результаты в первичных образцах пациентов, мы выполнили анализ дифференциальной экспрессии генов в образцах с (n = 22) и без (n = 216) мутациями BCOR из исследования Beat AML (Tyner et al., 2018). Дифференциально экспрессируемые гены в образцах с мутацией BCOR были обогащены регуляторами плюрипотентности стволовых клеток и сигнальных путей рака с более высокими уровнями экспрессии генов FGFR1, PDGFA и FOXO1 (рисунки 6C, 6D, S5C и S5D), аналогично тому, что мы наблюдали в BCOR. мутировавшие модели клеточной линии.
Используя анализ ChIP-seq и RNA-seq h4K27me3 и h3AK119ub, мы обнаружили, что гены-мишени PRC1.1 JAG1 и FOXO1 имели репрессированное состояние хроматина и слабо экспрессировались в первичных AML-бластах BCOR WT , тогда как эти гены были связаны с активным состоянием хроматина в первичных бластах BCOR PUFD-Tr AML (Фигуры 6E и S5E). В целом эти данные указывают на то, что инактивация PRC1.1 в AML с мутацией BCOR приводит к аберрантной экспрессии сигнальных программ стволовых клеток через эпигенетическую дерепрессию генов-мишеней.
BCOR KO вызывает частичное восстановление PRC1.1 и активирует набор высокочувствительных генов-мишенейУ пациентов с миелоидными злокачественными новообразованиями мутации BCOR чаще всего обнаруживаются без одновременных мутаций BCORL1, что указывает на то, что частичное нарушение PRC1.1 является достаточным для запуска клонального преимущество. Мы предположили, что подмножество функционально значимых генов-мишеней PRC1.1 особенно чувствительно к количественному снижению численности PRC1.1. Поэтому мы сравнили влияние условий одиночного BCOR PUFD-Tr и BCOR KO на экспрессию генов с действием условий двойного мутанта (BCOR PUFD-Tr / BCORL1 KO и BCOR KO / BCORL1 KO ) и обнаружили, что подмножество генов, которые были активированы в условиях двойного мутанта BCOR / BCORL1, также индуцировались в одиночных мутантах BCOR.(Рисунок 7A и S6A).
biorxiv; 2021.03.08.433705v1 / FIG7F7fig7 Рисунок 7. Мутации(A) Диаграммы Венна представляют собой перекрытия значительно активированных генов в мутантных клетках PRC1.1 по сравнению с контролем WT.
(B) Графики в виде прямоугольников и усов, представляющие уровни экспрессии (log2-преобразованные нормализованные значения DESeq2) генов C1 (n = 65) и C2 (n = 567) в WT, BCORL1 KO , BCOR KO или BCOR / BCORL1 КО ячеек.Гены C1 и C2 были определены с использованием неконтролируемого иерархического кластерного анализа уровней экспрессии PRC1.1-чувствительных генов в мутантных клетках WT или PRC1.1, показанных на рисунке 5C. Линия в середине прямоугольника представляет собой медианное значение, края прямоугольника представляют 25-й и 75-й процентили, а усы показывают 5-й и 95-й процентили. Для статистического анализа использовался критерий суммы рангов Вилкоксона. n.s. р> 0,05, **** р <0,0001.
(C) График вулкана, представляющий изменения экспрессии генов с усиленной регуляцией в BCOR KO vs.WT (левая панель) и клетки BCOR KO / BCORL1 KO по сравнению с клетками WT (правая панель) K562. Показаны только PRC1.1-чувствительные гены кластера C1 (n = 65). Изменения кратности Log2 1 и padj 0,05 показаны пунктирными линиями.
(D) Данные о покрытии ChIP-seq и ATAC-seq отслеживают PRC1.1-чувствительный ген FGFR1 в WT или мутантных PRC1.1 клетках K562.
(E) Уровни экспрессии гена FGFR1 (нормализованное количество DESeq2) в клетках WT или PRC1.1 мутантных K562 (верхняя панель) представлены как среднее ± SEM.Соответствующий вестерн-блоттинг-анализ уровней белка FGFR1 (нижняя панель) (FGFR1: 91 кДа, гликозилированный FGFR1: 120 кДа).
(F) Схема экспериментального рабочего процесса для конкурентных анализов in vitro. 80% положительных контрольных клеток RFP657 смешивали с 20% BFP-положительных конкурентных клеток и культивировали в присутствии ДМСО или указанных лекарств с добавлением или без добавления FGF2 (10 нг / мл). Экспрессию флуорофора отслеживали с течением времени с помощью анализа проточной цитометрии. Конкурентные условия экспрессировали не нацеленный контроль или sgRNA, нацеленные на BCORL1, BCOR, BCOR и BCORL1 или KDM2B.
(G) Тепловые карты, представляющие процентное соотношение участников за 10 дней. Клетки культивировали в присутствии ДМСО (верхняя панель) или 1 мкМ иматиниба (нижняя панель) с добавлением или без добавления FGF2 (10 нг / мл).
(H) Процент конкурентных клеток, экспрессирующих sgRNAs, нацеленных на BCOR / BCORL1 (выделено красным) или не нацеливающий контроль (выделено темно-серым) с течением времени. Клетки обрабатывали только 1 мкМ иматиниба (сплошная линия) или 1 мкМ иматиниба и 1 мкМ FGFRi (PD173074) в комбинации (пунктирная линия).
В зависимости от их чувствительности к частичной инактивации PRC1.1, ответные гены сгруппированы в две группы: кластер 1 (C1, n = 65) и кластер 2 (C2, n = 567) (рис. 5D). Гены C1 показали значительно более высокие уровни экспрессии в клетках с одиночным BCOR KO и двойным BCOR KO / BCORL1 KO по сравнению с генами C2 и были значительно более высоко экспрессированы в двойных клетках BCOR KO / BCORL1 KO , чем в клетках одиночные клетки BCOR KO (Рисунок 7B).Таким образом, снижение PRC1.1 в единичных мутантных клетках BCOR достаточно для активации подмножества высокочувствительных генов-мишеней PRC1.1, тогда как полная потеря PRC1.1 в клетках BCOR KO / BCORL1 KO приводит к активации полного набора целевых генов. гены.
Внутри кластера генов C1 рецептор 1 фактора роста фибробластов (FGFR1) был одним из наиболее высоко регулируемых генов в частично или полностью дефицитных мутантных клетках PRC1.1 (Фигуры 7C и S6B). На исходном уровне локус FGFR1 имел репрессированное состояние хроматина, отмеченное низкой доступностью, обогащением h3AK119ub и h4K27me3 и отсутствием h4K27ac (рис. 7D).Соответственно, не было транскрипции FGFR1 и детектируемого белка FGFR1 (фиг. 7E). Частичная инактивация PRC1.1 в клетках BCOR KO и BCOR PUFD-Tr вызвала значительно повышенную экспрессию FGFR1 без эпигенетического репрограммирования, тогда как полное нарушение PRC1.1 в BCOR KO / BCORL1 KO , BCOR PUFD-Tr / BCORL1 KO , PCGF1 KO и KDM2B KO вызвали потерю репрессивных гистоновых меток и повышение доступности хроматина, что коррелировало со значительным увеличением экспрессии гена FGFR1 и уровней белка (рисунки 7D и 7E).Чтобы подтвердить это наблюдение на независимой модели линии клеток AML, мы генерировали клетки MOLM13 с BCOR KO , BCORL1 KO , BCOR KO / BCORL1 KO или KDM2B KO , и обнаружили, что инактивация PRC1.1 вызывает аналогичная активация экспрессии FGFR1 (рисунок S6C). Чтобы определить, был ли эффект инактивации PRC1.1 на экспрессию FGFR1 обратимым и поддерживал ли он зависимость от интактного BCOR / PRC1.1, мы экспрессировали sgRNA-устойчивую полноразмерную кДНК BCOR в клетках BCOR KO / BCORL1 KO .Комплементация экспрессии BCOR восстанавливала сборку комплекса PRC1.1 (рисунок S2G) и снижала уровни белка FGFR1 в клетках с дефицитом PRC1.1 (рисунок S6D).
biorxiv; 2021.03.08.433705v1 / FIGS6F13figs6 Рисунок S6. Мутации(A) Число дифференциально экспрессируемых генов в мутанте PRC1.1 по сравнению с клетками WT K562 (кратное изменение> 2, padj <0,05). Дифференциально экспрессируемые гены были подразделены на основе их состояний хроматина, определенных на фиг. 5A.
(B) Графики вулкана, представляющие изменения экспрессии генов с усиленной регуляцией генов в мутанте PRC1.1 по сравнению с клетками WT K562. Показаны только PRC1.1-чувствительные гены кластера C1 (n = 65). Изменения кратности Log2 1 и padj 0,05 показаны пунктирными линиями.
(C) Вестерн-блот-анализ уровней белка FGFR1 в WT, BCORL1 KO , BCOR KO , BCOR KO / BCORL1 KO или KDM2B KO MOLM13 гликозилированных клеточных линиях AMLa1: FGFR1: 120 кДа).PRC1.1 мутантные клетки MOLM13 представляют собой основные популяции. Высокие частоты инделирования (> 90%) в соответствующих генах были подтверждены секвенированием по Сэнгеру и анализом TIDE (Brinkman et al., 2014).
(D) Вестерн-блот-анализ уровней белка FGFR1 в клетках WT, BCOR KO / BCORL1 KO (DKO) или BCOR KO / BCORL1 KO K562, дополненных клетками WT-BCOR (DKO16 ) BCOR . Экзогенный полноразмерный WT-BCOR стабильно экспрессировался в клетках DKO BCOR с помощью лентивирусной трансдукции.
(E) Частота Indel через 19 дней после конкуренции in vitro между контролем и клетками-конкурентами в присутствии 1 мкМ иматиниба и лиганда FGF2 (10 нг / мл), определенная секвенированием по Сэнгеру и анализом TIDE (Brinkman et al., 2014).
(F) Вестерн-блот-анализ уровней белка FGFR1 через 13 дней после конкуренции контрольных клеток с клетками-конкурентами in vitro в присутствии 1 мкМ иматиниба и лиганда FGF2 (10 нг / мл). FGFR1: 91 кДа, гликозилированный FGFR1: 120 кДа, 140 кДа.
(G) Контроль или BCOR КО / BCORL1 КО клетки обрабатывали ДМСО или ингибитором FGFR PD173074 в возрастающих концентрациях в течение 72 часов.Жизнеспособность клеток измеряли с помощью люминесцентного анализа CellTiter-Glo (повторений n = 2, полосы ошибок указывают стандартное отклонение (SD)).
Сообщается, что усиление экспрессии FGFR1 является механизмом устойчивости к ингибированию тирозинкиназы (Javidi-Sharifi et al., 2019; Traer et al., 2014 , 2016), а мутации BCOR были специфически связаны с бластной фазой ХМЛ и устойчивостью к ингибиторам, направленным на BCR-ABL (Branford et al., 2018). Поэтому мы предположили, что нарушение PRC1.1 вызывает устойчивость к целевому ингибированию BCR-ABL в клетках K562 посредством дерепрессии FGFR1. Чтобы проверить эту гипотезу, мы оценили относительный рост клеток с дефицитом PRC1.1 по сравнению с контрольными клетками с использованием конкурентных анализов in vitro. Мы смешали меченые флуорохромом контрольные клетки (Cas9 плюс не нацеливающую sgRNA) и клетки-конкуренты (Cas9 плюс sgRNA, нацеленную на BCOR, BCORL1, BCOR / BCORL1 или KDM2B) в соотношении 80:20, затем использовали проточную цитометрию для измерения их относительной доли. более 10 дней с добавлением ингибиторов или без них (рис. 7F).В исходных условиях (среда с 10% сыворотки, ДМСО) клетки с мутацией PRC1.1 не обладали преимуществом селективного роста по сравнению с контрольными клетками (фигура 7G). Напротив, когда мы заблокировали передачу сигналов BCR-ABL с помощью иматиниба (1 мкМ), клетки BCOR KO , BCOR KO / BCORL1 KO и KDM2B KO показали преимущество избирательного роста по сравнению с контрольными клетками, что указывает на повышенную относительную устойчивость. к иматинибу (рис. 7G). Одиночные клетки BCOR KO , которые показали менее выраженную активацию сигнальных генов, включая FGFR1 (рисунки 7C и 7E), были менее устойчивы к иматинибу, чем двойные клетки BCOR KO / BCORL1 KO или KDM2B KO , что отражается более низкой долей BFP-положительных клеток на 10 день (BCOR KO : 27.5%; BCOR KO / BCORL1 KO : 52,1%; KDM2B KO : 49,7%) (Рисунок 7G). В каждом конкурентном состоянии мы наблюдали сопутствующее увеличение доли инделя на сайтах-мишенях sgRNA, подтверждая корреляцию между флуоресцентными маркерами и направлением генов (рисунок S6E).
Чтобы определить, опосредован ли эффект инактивации PRC1.1 на устойчивость к иматинибу активностью FGFR1, мы измерили функциональные последствия увеличения или ингибирования передачи сигналов FGFR1.
Добавление среды с FGF2 (10 нг / мл) увеличивало величину избирательного преимущества клеток BCOR KO , BCOR KO / BCORL1 KO и KDM2B KO по сравнению с контрольными клетками (рис. 7G) и коррелировало с высокими уровнями белка FGFR1 на 13 день (рисунок S6F).Напротив, одновременная обработка клеток иматинибом и селективным низкомолекулярным ингибитором FGFR1 PD173074 (10 нг / мл) полностью устраняла устойчивость клеток BCOR KO / BCORL1 KO по сравнению с клетками BCOR KO / BCORL1 KO , обработанными клетками только иматиниб (фигура 7H). Обработка клеток K562 одним PD173074 не влияла на рост или выживаемость клеток (фиг. S6G). Вместе эти данные указывают на то, что генетическая инактивация PRC1.1 вызывает устойчивость к иматинибу через его эффекты на FGFR1.
ОбсуждениеЗдесь мы объединили генетический анализ 6162 первичных образцов пациентов с функциональными исследованиями клеточной линии и первичных образцов AML, чтобы определить роль изменений PRC1.1 в патогенезе миелоидных злокачественных новообразований. Мы обнаружили, что BCOR и BCORL1 являются центральными адаптерами, которые необходимы для сборки комплекса PRC1.1 и репрессии целевого гена. Связанные с лейкемией мутации BCOR и BCORL1 взаимодействуют, чтобы управлять прогрессированием заболевания, избирательно отделяя ферментативное ядро RING / PCGF1 от комплекса, тем самым вызывая количественное снижение ферментативно активного PRC1.1 на хроматине и приводя к аберрантной активации онкогенных сигнальных программ. Далее мы демонстрируем, что дерепрессия генов-мишеней PRC1.1 в клетках лейкемии с мутациями BCOR / BCORL1 опосредует функциональную устойчивость к лечению ингибиторами киназ, которая обратима с ингибированием FGFR1.
Мутации онкогенных генов, которые вводят кодон преждевременной терминации, могут действовать по множеству механизмов, включая потерю или усиление функции в зависимости от экспрессируемого белкового продукта (Lindeboom et al., 2019; Ян и др., 2018). Здесь мы показываем, что патогенные мутации BCOR имеют различные последствия для экспрессии белка BCOR, но имеют общий механизм их эпигенетических и онкогенных эффектов. В частности, все мутации BCOR, связанные с лейкемией, вызывают потерю стабилизирующего взаимодействия между C-концевым доменом BCOR-PUFD и PRC1.1-специфическим адаптером PCGF1, что приводит к отделению ферментативного ядра PRC1 от PRC1, нацеленного на хроматин. 1 вспомогательный подкомплекс. Эта парадигма иллюстрируется подмножеством мутаций BCOR у пациентов, которые вызывают стабильную экспрессию усеченных PUFD белков, избегая нонсенс-опосредованного распада.Мы показываем как на образцах пациентов, так и на моделях сконструированных клеточных линий, что эти усекающие PUFD мутации BCOR нарушают связывание и стабильность PCGF1 и приводят к дерепрессии основного набора мишеней PRC1.1. Кроме того, мы обнаружили, что экспрессия BCOR-усеченного PUFD была функционально эквивалентна полной потере BCOR, что указывает на то, что изолированное нарушение взаимодействия белок-белок BCOR-PCGF1 является достаточным для повышения патогенности миелоидных злокачественных новообразований. Примечательно, что C-концевой усеченный BCOR, который потерял связывание с PCGF1, все еще был способен взаимодействовать с KDM2B, в соответствии с недавним исследованием, показывающим, что экзогенно экспрессируемый BCOR без PUFD связывает KDM2B, но не PCGF1 (Wang et al., 2018). Образование стабильного субкомплекса KDM2B-BCOR в отсутствие связывания PCGF1 может иметь более широкие последствия для онкогенеза. Например, предполагается, что внутренние тандемные дупликации (ITD) в доменах BCOR PUFD у пациентов со светлоклеточной саркомой почки (CCSK), CNS-PNET и другими видами рака нарушают взаимодействие PCGF1 (Wong et al., 2020). Мы предполагаем, что потеря PCGF1 может позволить онкобелкам захватить субкомплексы BCOR-ITD-KDM2B, что приводит к усилению или изменению функции PRC1.1, обеспечивая концептуальную модель, с помощью которой PRC1.1 изменения могут управлять онкогенезом в специфическом контексте рака.
Наше исследование обеспечивает механистическую основу для более высокой частоты мутаций BCOR при лейкемии по сравнению с BCORL1. Мы обнаружили, что BCOR и BCORL1 участвуют в функционально избыточных и взаимоисключающих видах PRC1.1, разделяя ключевые партнеры по взаимодействию с белками и располагаясь в одном и том же наборе локусов хроматина. Таким образом, совокупная активность PRC1.1 в клетке, по-видимому, отражает совокупность активности комплекса BCOR-PRC1.1 и BCORL1-PRC1.1.Далее мы обнаружили, что BCOR был значительно выше экспрессирован, чем BCORL1 при нормальном и злокачественном гематопоэзе, и что мутации BCOR не приводили к компенсаторному увеличению уровней экспрессии гена BCORL1 или наоборот. Следовательно, мутации BCOR вызывали более выраженное количественное снижение глобальных уровней PRC1.1, чем мутации BCORL1, которые коррелировали с активацией транскрипции набора высокочувствительных генов-мишеней PRC1.1. Наши данные показывают, что мутации BCOR и BCORL1 взаимодействуют путем прогрессивного снижения PRC1.1 и дозозависимая дерепрессия генов-мишеней PRC1.1. В соответствии с этой моделью, мутации BCORL1 обычно возникали в контексте ранее существовавшей мутации BCOR у пациентов с миелоидными злокачественными новообразованиями, которые были связаны с прогрессированием заболевания.
Мы обнаружили, что PCGF1 или KDM2B редко мутировали у пациентов с AML, но эта двуаллельная генетическая инактивация PCGF1 или KDM2B в нашей модели клеточной линии AML имитировала функциональные последствия мутаций BCOR / BCORL1. BCOR и BCORL1 расположены на X-хромосоме и, как сообщается, не избегают X-инактивации при раке (Grossmann et al., 2011), указывая на то, что полное функциональное нарушение BCOR или BCORL1 может быть достигнуто с помощью одного мутационного события у людей с генотипом XX или XY. Напротив, PCGF1 (хромосома 2) и KDM2B (хромосома 12) являются аутосомными генами и потребуют двух мутационных событий, чтобы вызвать дерепрессию целевого гена, что приведет к аналогичному клональному преимуществу. В соответствии с этим наблюдением, единственная мутация PCGF1, которую мы идентифицировали в когорте AML, присутствовала во фракции аллелей с высокой вариативностью, что указывает на потерю гетерозиготности.
Предыдущие исследования показали, что отложение h3AK119ub неканоническими комплексами PRC1 было критическим для репрессии целевого гена и рекрутирования PRC2 (Blackledge et al., 2020; Fursova et al., 2019; Tamburri et al., 2020). Это согласуется с нашим открытием, что нарушение PRC1.1 приводит к потере h3AK119ub и снижению связывания PRC2 в генах-мишенях, что коррелирует с активацией транскрипции. В то время как мы наблюдали широко распространенное связывание PRC1.1 с репрессированными промоторами, только субнабор PRC1, богатого CpG.Связанные гены 1 были дерепрессированы при разрыве PRC1.1. Ранее сообщалось о синергических, а также об уникальных функциях неканонических комплексов PRC1, предполагающих, что комплексы PRC1.3 / 5 или PRC1.6 могут компенсировать потерю PRC1.1 в общих генах-мишенях (Scelfo et al., 2019; Tamburri et al. ., 2020). Недавнее исследование показало, что комплексы PRC1 способствуют активной экспрессии генов в эмбриональных предшественниках эпидермиса мыши (Cohen et al., 2018). Хотя мы наблюдали связывание PRC1.1 с активными промоторами генов при лейкемии, мы не обнаружили изменений в уровнях экспрессии активных генов на PRC1.1 срыв. Поэтому мы предпочитаем модель, предложенную Farcas et al. (Farcas et al., 2012), в которых PRC1.1 может пробовать промоторы генов, богатых CpG, на чувствительность к Polycomb-опосредованному молчанию независимо от состояния их хроматина, тогда как дополнительные факторы необходимы для облегчения репрессии генов.
Идентификация специфических генных мутаций или сигнатур экспрессии генов может определять лекарственные реакции у пациентов с миелоидными злокачественными новообразованиями (Tyner et al., 2018). Мы обнаружили, что как в моделях клеточных линий, так и в образцах первичного AML, PRC1.1 был связан с активацией клеточных сигнальных путей, регулирующих плюрипотентность стволовых клеток, что согласуется с ранее предполагаемой ролью PRC1.1 в регуляции программ транскрипции стволовых клеток (Kelly et al., 2019; Wang et al., 2018). Мы также продемонстрировали, что эпигенетическая активация FGFR1 гена-мишени PRC1.1 опосредует устойчивость к иматинибу в PRC1.1-дефицитных клеточных линиях K562, которую можно преодолеть с помощью ингибирования FGFR1. Наши данные показывают, что клетки с дефицитом PRC1.1 были способны поддерживать передачу сигналов и выживаемость клеток в отсутствие передачи сигналов BCR-ABL за счет использования FGFR1 в качестве альтернативного пути передачи сигналов.Это подтверждается предыдущими исследованиями, демонстрирующими, что активация передачи сигналов FGFR была механизмом устойчивости к TKI в клетках K562 (Traer et al., 2014) и что мутации BCOR и BCORL1 были связаны с бластным кризисом у пациентов с CML (Branford et al., 2018). ). Наши результаты также предполагают, что мутации BCOR и BCORL1 могут в более широком смысле определять группу миелоидных злокачественных новообразований с дефицитом PRC1.1, которые отвечают на фармакологическое ингибирование гиперактивной передачи сигналов RAS / MAPK.
Globine — Kaj pa človekove pravice?
Доктор.Андрей Сая | (foto: Katoliška Cerkev)
13.04.2021, 19:10 Уредништво Радия Огнище
V Globinah ostajamo pri papeževi okrožnici Vsi bratje.Tokrat smo se lotili vprašanja človekovih pravic, ki se sistematično kršijo na all koncih sveta, po other strani pa se nanje sklicujemo pogosto takrat, ko nam ustreza zaradi lastnih interesov.
Гре за защиту чловека кот посамезника, за наук, да те права ижаяо из чловекове нараве. Ko je človek rojen, te pravice prinese na svet.Не подели му джи нобен краль, нобена влада в нобен председник. Beseda je v nadaljavanju tekla tudi o krščanskem pogledu na človekove pravice nekoč in danes, o kršitvah в различных интерпретациях тега, kar naj bi bilo temeljno in univerzalno za vse človeštas. Kakšne pravice imajo migranti in ali je odločitev za oziroma proti cepljenju tudi človekova pravica?
Z nami je bil profesor cerkvenega prava dr.