
Законы действия постоянного тока на возбудимые ткани
Полярный закон действия тока. При раздражении нерва или мышцы постоянным током возбуждение возникает в момент замыкания постоянного тока только под катодом, а в момент размыкания — только под анодом, причем порог замыкательного удара меньше, чем размыкательного. Прямые измерения показали, что прохождение через нервное или мышечное волокно электрического тока вызывает прежде всего изменение мембранного потенциала под электродами. В области приложения к поверхности ткани анода (+) положительный потенциал на наружной поверхности мембраны возрастает, т.е. в этом участке происходит гиперполяризация мембраны, что не способствует возбуждению, а, наоборот, ему препятствует. В том же участке, где к мембране приложен катод (-), положительный потенциал наружной поверхности снижается, происходит деполяризация, и если она достигает критической величины — в этом месте возникает ПД.
Изменение МП возникают не только непосредственно в точках приложения к нервному волокну катода и анода, но и на некотором расстоянии от них, но величина этих сдвигов убывает по мере удаления от электродов. Изменения МП под электродами носят название электротонических ( соответственно кат-электротон и ан-электротон), а за электродами — периэлектротонических (кат- и ан-периэлектротон).
Изменения МП под электродами носят название электротонических ( соответственно кат-электротон и ан-электротон), а за электродами — периэлектротонических (кат- и ан-периэлектротон).
Увеличение МП под анодом (пассивная гиперполяризация) не сопровождается изменением ионной проницаемости мембраны даже при большой силе приложенного тока. Поэтому при замыкании постоянного тока возбуждение под анодом не возникает. В отличие от этого, уменьшение МП под катодом (пассивная деполяризация) влечет за собой кратковременное повышение проницаемости для Na, что приводит к возбуждению.
Повышение проницаемости мембраны для Na при пороговом раздражении не сразу достигает максимальной величины. В первый момент деполяризация мембраны под катодом приводит к небольшому увеличению натриевой проницаемости и открытию небольшого числа каналов. Когда же под влиянием этого в протоплазму начинают поступать заряженные положительно ионы Na+, то деполяризация мембраны усиливается. Это ведет к открытию других Na-каналов, и, следовательно, к дальнейшей деполяризации, которая, в свою очередь, обуславливает еще большее повышение натриевой проницаемости. Этот круговой процесс, основанный на т.н. положительной обратной связи, получил название регенеративной деполяризации. Возникает она только при снижении Ео до критического уровня (Ек ). Причина повышения натриевой проницаемости при деполяризации связана, вероятно, с удалением Са++ из натриевых ворот при возникновении электро отрицательности (или снижении электро положительности) на наружной стороне мембраны.
Этот круговой процесс, основанный на т.н. положительной обратной связи, получил название регенеративной деполяризации. Возникает она только при снижении Ео до критического уровня (Ек ). Причина повышения натриевой проницаемости при деполяризации связана, вероятно, с удалением Са++ из натриевых ворот при возникновении электро отрицательности (или снижении электро положительности) на наружной стороне мембраны.
Повышенная натриевая проницаемость через десятые доли миллисекунды за счет механизмов натриевой инактивации прекращается.
Скорость, с которой происходит деполяризация мембраны, зависит от силы раздражающего тока. При слабой силе деполяризация развивается медленно, и поэтому для возникновения ПД такой стимул должен иметь большую длительность.
Локальный ответ, который возникает при под пороговых стимулах, так же, как и ПД, обусловлен повышением натриевой проницаемости мембраны. Однако при под пороговом стимуле это повышение недостаточно велико для того, чтобы вызвать процесс регенеративной деполяризации мембраны. Поэтому начавшаяся деполяризация приостанавливается инактивацией и повышением калиевой проницаемости.
Поэтому начавшаяся деполяризация приостанавливается инактивацией и повышением калиевой проницаемости.
Подводя итог изложенному выше, можно следующим образом изобразить цепь событий, развивающихся в нервном или мышечном волокне под катодом раздражающего тока: пассивная деполяризация мембраны —- повышение натриевой проницаемости — усиление потока Na внутрь волокна — активная деполяризация мембраны — локальный ответ — превышение Ек — регенеративная деполяризация — потенциал действия (ПД).
Каков же механизм возникновения возбуждения под анодом при размыкании? В момент включения тока под анодом мембранный потенциал возрастает — происходит гиперполяризация. При этом разница между Ео и Ек растет, и для того, чтобы сдвинуть МП до критического уровня, нужна большая сила. При выключении тока (размыкание) исходный уровень Ео восстанавливается. Казалось бы, в это время нет условий для возникновения возбуждения. Но это справедливо только для того случая, если действие тока продолжалось очень короткое время (менее 100 мсек.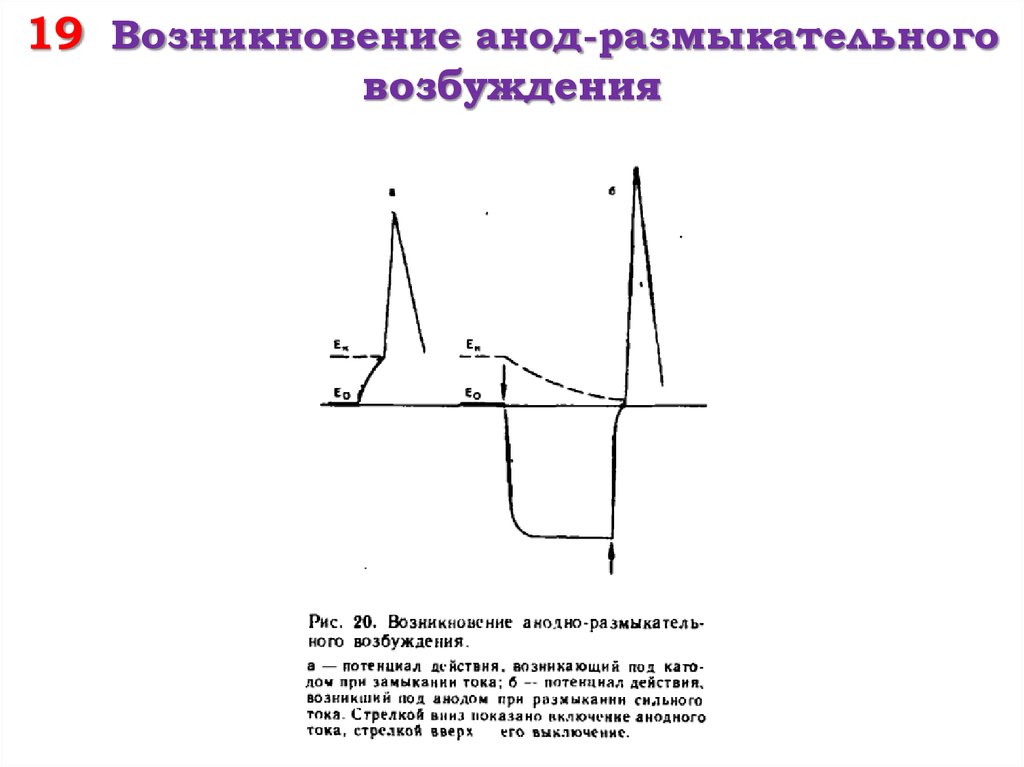 ). При длительном действии тока начинает меняться сам критический уровень деполяризации — он растет. И, наконец, возникает момент, когда новый Ек становится равным старому уровню Ео. Теперь при выключении тока возникают условия для возбуждения, ибо мембранный потенциал становится равным новому критическому уровню деполяризации. Величина ПД при размыкании всегда больше, чем при замыкании.
). При длительном действии тока начинает меняться сам критический уровень деполяризации — он растет. И, наконец, возникает момент, когда новый Ек становится равным старому уровню Ео. Теперь при выключении тока возникают условия для возбуждения, ибо мембранный потенциал становится равным новому критическому уровню деполяризации. Величина ПД при размыкании всегда больше, чем при замыкании.
Зависимость пороговой силы стимула от его длительности. Как уже указывалось, пороговая сила любого стимула в определенных пределах находится в обратной зависимости от его длительности. В особенно четкой форме эта зависимость проявляется при использовании в качестве раздражителя прямоугольных толчков постоянного тока. Полученная в таких опытах кривая получила название «кривой силы-времени». Она была изучена Гоорвегом, Вейсом и Лапиком в начале века. Из рассмотрения этой кривой прежде всего следует, что ток ниже некоторой минимальной величины или напряжения не вызывает возбуждения, как бы длительно он не действовал. Минимальная сила тока, способная вызвать возбуждение, названа Лапиком реобазой. Наименьшее время, в течение которого должен действовать раздражающий стимул, называют полезным временем. Усиление тока приводит к укорочению минимального времени раздражения, но не беспредельно. При очень коротких стимулах кривая силы-времени становится параллельной оси координат. Это означает, что при таких кратковременных раздражениях возбуждения не возникает, как бы ни была велика сила раздражения.
Минимальная сила тока, способная вызвать возбуждение, названа Лапиком реобазой. Наименьшее время, в течение которого должен действовать раздражающий стимул, называют полезным временем. Усиление тока приводит к укорочению минимального времени раздражения, но не беспредельно. При очень коротких стимулах кривая силы-времени становится параллельной оси координат. Это означает, что при таких кратковременных раздражениях возбуждения не возникает, как бы ни была велика сила раздражения.
Определение полезного времени практически затруднено, так как точка полезного времени находится на участке кривой, переходящей в параллельную. Поэтому Лапик предложил использовать полезное время двух реобаз — хронаксию. Ее точка находится на самом крутом участке кривой Гоорвега-Вейса. Хронаксиметрия получила широкое распространение как в эксперименте, так и в клинике для диагностики повреждений волокон двигательных нервов.
Зависимость порога от крутизны нарастания силы раздражителя. Величина порога раздражения нерва или мышцы зависит не только от длительности стимула, но и от крутизны нарастания его силы. Порог раздражения имеет наименьшую величину при толчках тока прямоугольной формы, характеризующихся максимально быстрым нарастанием тока. Если же вместо таких стимулов применять линейно или экспоненциально нарастающие стимулы, пороги оказываются увеличенными и тем больше, чем медленнее нарастает ток. При уменьшении крутизны нарастания тока ниже некоторой минимальной величины (т.н. критический наклон) ПД вообще не возникает, до какой бы конечной силы не увеличивался ток.
Величина порога раздражения нерва или мышцы зависит не только от длительности стимула, но и от крутизны нарастания его силы. Порог раздражения имеет наименьшую величину при толчках тока прямоугольной формы, характеризующихся максимально быстрым нарастанием тока. Если же вместо таких стимулов применять линейно или экспоненциально нарастающие стимулы, пороги оказываются увеличенными и тем больше, чем медленнее нарастает ток. При уменьшении крутизны нарастания тока ниже некоторой минимальной величины (т.н. критический наклон) ПД вообще не возникает, до какой бы конечной силы не увеличивался ток.
Такое явление приспособления возбудимой ткани к медленно нарастающему раздражителю получило название аккомодация. Чем выше скорость аккомодации, тем более круто должен нарастать стимул, чтобы не утратить своего раздражающего действия. Аккомодация к медленно нарастающему току обусловлена тем, что за время действия этого тока в мембране успевают развиться процессы, препятствующие возникновению ПД.
Выше уже указывалось, что деполяризация мембраны приводит к началу двух процессов: одного — быстрого, ведущего к повышению натриевой проницаемости и возникновению ПД, а другого — медленного, приводящего к инактивации натриевой проницаемости и к окончанию возбуждения. При крутом нарастании стимула Na-активация успевает достичь значительной величины прежде, чем развивается Na-инактивация. В случае медленного нарастания силы тока на первый план выступают процессы инактивации, приводящие к повышению порога и снижению амплитуды ПД. Все агенты, усиливающие или ускоряющие инактивацию, увеличивают скорость аккомодации.
Аккомодация развивается не только при раздражении возбудимых тканей электрическим током, но и в случае применения механических, термических и прочих стимулов. Так, быстрый удар палочкой по нерву вызывает его возбуждение, при медленном же надавливании на нерв той же палочкой возбуждения не возникает. Изолированное нервное волокно можно возбудить быстрым охлаждением, а медленным — нельзя. Лягушка выпрыгнет, если ее бросить в воду с температурой 40 градусов, но если ту же лягушку поместить в холодную воду, и медленно нагревать ее, то животное сварится, но не будет реагировать прыжком на подъем температуры.
Лягушка выпрыгнет, если ее бросить в воду с температурой 40 градусов, но если ту же лягушку поместить в холодную воду, и медленно нагревать ее, то животное сварится, но не будет реагировать прыжком на подъем температуры.
В лаборатории показателем скорости аккомодации является та наименьшая крутизна нарастания тока, при которой стимул еще сохраняет способность вызывать ПД. Эту минимальную крутизну называют критическим наклоном. Его выражают или в абсолютных единицах (мА/сек), или в относительных (как отношение пороговой силы того постепенно нарастающего тока, который еще способен вызывать возбуждение, к реобазе прямоугольного толчка тока).
Закон «все или ничего». При изучении зависимости эффектов раздражения от силы приложенного стимула был установлен т.н. закон «все или ничего». Согласно этому закону, под пороговые раздражения не вызывают возбуждения («ничего»), при пороговых же стимулах возбуждение сразу приобретает максимальную величину («все»), и уже не возрастает при дальнейшем усилении раздражителя.
Эта закономерность первоначально была открыта Боудичем при исследовании сердца, а в дальнейшем подтверждена и на других возбудимых тканях. Долгое время закон «все или ничего» неправильно интерпретировался как общий принцип реагирования возбудимых тканей. Предполагали, что «ничего» означает полное отсутствие ответа на под пороговый стимул, а «все» рассматривалось как проявление полного исчерпания возбудимым субстратом его потенциальных возможностей. Дальнейшие исследования, в особенности микроэлектродные, показали, что эта точка зрения не соответствует действительности. Выяснилось, что при под пороговых силах возникает местное не распространяющееся возбуждение (локальный ответ). Вместе с тем оказалось, что «все» также не характеризует того максимума, которого может достигнуть ПД. В живой клетке существуют процессы, активно приостанавливающие деполяризацию мембраны. Если каким-либо воздействием на нервное волокно, например, наркотиками, ядами, ослабить входящий Na-ток, обеспечивающий генерацию ПД, то он перестает подчиняться правилу «все или ничего» — его амплитуда начинает градуально зависеть от силы стимула. Поэтому «все или ничего» рассматривается сейчас не как всеобщий закон реагирования возбудимого субстрата на раздражитель, а лишь как правило, характеризующее особенности возникновения ПД в данных конкретных условиях.
Поэтому «все или ничего» рассматривается сейчас не как всеобщий закон реагирования возбудимого субстрата на раздражитель, а лишь как правило, характеризующее особенности возникновения ПД в данных конкретных условиях.
Понятие о возбудимости. Изменения возбудимости при возбуждении.
Параметры возбудимости.
Возбудимостью называется способность нервной или мышечной клетки отвечать на раздражение генерацией ПД. Основным мерилом возбудимости обычно служит реобаза. Чем она ниже, тем выше возбудимость, и наоборот. Связано это с тем, что, как мы уже говорили ранее, главным условием возникновения возбуждения является достижение МП критического уровня деполяризации (Ео <= Ек). Поэтому мерилом возбудимости является разница между этими величинами (Ео — Ек). Чем меньше эта разница, тем меньшую силу надо приложить к клетке, чтобы сдвинуть мембранный потенциал до критического уровня, и, следовательно, тем больше возбудимость клетки.
Еще Пфлюгер показал, что возбудимость — величина непостоянная. Катод повышает возбудимость, анод ее понижает. Напомним, что эти изменения возбудимости под электродами носят название электротонических. Русский ученый Вериго показал, что при длительном действии постоянного тока на ткань, или при действии сильных раздражителей эти электротонические изменения возбудимости извращаются — под катодом начальное повышение возбудимости сменяется ее понижением (развивается т.н. катодическая депрессия), а под анодом сниженная возбудимость постепенно возрастает. Причина этих изменений возбудимости на полюсах постоянного тока связана с тем, что величина Ек меняется при длительном действии раздражителя. Под катодом (и при возбуждении) Ек постепенно удаляется от МП, снижается, так что наступает момент, когда разница Е0-Ек становиться больше исходной. Это приводит к падению возбудимости ткани. Напротив, под анодом Ек имеет тенденцию к возрастанию, постепенно приближаясь к Ео. Возбудимость при этом растет, так как уменьшается исходная разница между Ео и Ек.
Причиной изменения критического уровня деполяризации под катодом является инактивация натриевой проницаемости, обусловленная длительной деполяризацией мембраны. Вместе с этим значительно повышается проницаемость для К. Все это приводит к тому, что мембрана клетки утрачивает способность отвечать на действие раздражающих стимулов. Такие же изменения мембраны лежат и в основе рассмотренного уже явления аккомодации. Под анодом же при действии тока явления инактивации снижаются .
Изменения возбудимости при возбуждении. Возникновение в нервном или мышечном волокне ПД сопровождается многофазными изменениями возбудимости. Для их изучения нерв или мышцу подвергают действию двух коротких электрических стимулов, следующих друг за другом с определенным интервалом. Первый называется раздражающим, второй — тестирующим. Регистрация возникающих в ответ на эти раздражения ПД позволила установить важные факты.
Во время локального ответа возбудимость повышена, так как мембрана деполяризована и разность между Е0 и Ек падает. Периоду же возникновения и развития пика потенциала действия соответствует полное исчезновение возбудимости, получившее название абсолютной рефрактерности (невпечатлительности). В это время тестирующий стимул не способен вызвать новый ПД, как бы сильно ни было это раздражение. Длительность абсолютной рефрактерности примерно совпадает с длительностью восходящей ветви ПД. В быстро проводящих нервных волокнах она составляет 0,4-0,7 мсек. В волокнах мышцы сердца — 250-300 мсек. Вслед за абсолютной рефрактерностью начинается фаза относительной рефрактерности , которая длится 4-8 мсек. Она совпадает с фазой реполяризации ПД. В это время возбудимость постепенно возвращается к первоначальному уровню. В этот период нервное волокно способно ответить на сильное раздражение, но амплитуда ПД будет резко снижена.
Периоду же возникновения и развития пика потенциала действия соответствует полное исчезновение возбудимости, получившее название абсолютной рефрактерности (невпечатлительности). В это время тестирующий стимул не способен вызвать новый ПД, как бы сильно ни было это раздражение. Длительность абсолютной рефрактерности примерно совпадает с длительностью восходящей ветви ПД. В быстро проводящих нервных волокнах она составляет 0,4-0,7 мсек. В волокнах мышцы сердца — 250-300 мсек. Вслед за абсолютной рефрактерностью начинается фаза относительной рефрактерности , которая длится 4-8 мсек. Она совпадает с фазой реполяризации ПД. В это время возбудимость постепенно возвращается к первоначальному уровню. В этот период нервное волокно способно ответить на сильное раздражение, но амплитуда ПД будет резко снижена.
Согласно ионной теории Ходжкина-Хаксли, абсолютная рефрактерность обусловлена вначале наличием максимальной натриевой проницаемости, когда новый стимул не может что-то изменить или добавить, а затем развитием натриевой инактивации, закрывающей Na-каналы. Вслед за этим происходит снижение натриевой инактивации, в результате чего постепенно восстанавливается способность волокна генерировать ПД. Это — состояние относительной рефрактерности.
Вслед за этим происходит снижение натриевой инактивации, в результате чего постепенно восстанавливается способность волокна генерировать ПД. Это — состояние относительной рефрактерности.
Относительная рефрактерная фаза сменяется фазой повышенной (супернормальной) возбудимости, совпадающей по времени с периодом следовой деполяризации. В это время разность между Ео и Ек ниже исходной. В двигательных нервных волокнах теплокровных животных длительность супернормальной фазы составляет 12-30 мсек.
Период повышенной возбудимости сменяется субнормальной фазой , которая совпадает со следовой гиперполяризацией. В это время разница между мембранным потенциалом (Ео) и критическим уровнем деполяризации (Ек) увеличивается. Длительность этой фазы составляет несколько десятков или сотен мсек.
Лабильность. Мы рассмотрели основные механизмы возникновения и распространения в нервных и мышечных волокнах одиночной волны возбуждения. Однако в естественных условиях существования организма по нервным волокнам проходят не одиночные, а ритмические залпы потенциалов действия. В чувствительных нервных окончаниях, расположенных в любых тканях, возникают и распространяются по отходящим от них афферентным нервным волокнам ритмические разряды импульсов даже при очень кратковременном раздражении. Равным образом из ЦНС по эфферентным нервам идет поток импульсов на периферию к исполнительным органам. Если исполнительным органом являются скелетные мышцы, то в них возникают вспышки возбуждений в ритме поступающих по нерву импульсов.
Однако в естественных условиях существования организма по нервным волокнам проходят не одиночные, а ритмические залпы потенциалов действия. В чувствительных нервных окончаниях, расположенных в любых тканях, возникают и распространяются по отходящим от них афферентным нервным волокнам ритмические разряды импульсов даже при очень кратковременном раздражении. Равным образом из ЦНС по эфферентным нервам идет поток импульсов на периферию к исполнительным органам. Если исполнительным органом являются скелетные мышцы, то в них возникают вспышки возбуждений в ритме поступающих по нерву импульсов.
Частота разрядов импульсов в возбудимых тканях может варьировать в широких пределах в зависимости от силы приложенного раздражения, свойств и состояния ткани и от скорости протекания отдельных актов возбуждения в ритмическом ряду. Для характеристики этой скорости Н.Е.Введенским и было сформулировано понятие лабильность. Под лабильностью, или функциональной подвижностью он понимал большую или меньшую скорость протекания тех элементарных реакций, которыми сопровождается возбуждение. Мерой лабильности является наибольшее число потенциалов действия, которое возбудимый субстрат способен воспроизвести в единицу времени в соответствии с частотой подаваемого раздражения.
Мерой лабильности является наибольшее число потенциалов действия, которое возбудимый субстрат способен воспроизвести в единицу времени в соответствии с частотой подаваемого раздражения.
Первоначально предполагали, что минимальный интервал между импульсами в ритмическом ряду должен соответствовать длительности абсолютного рефрактерного периода. Точные исследования, однако, показали, что при частоте следования стимулов с таким интервалом возникают только два импульса, а третий выпадает вследствие развивающейся депрессии. Поэтому интервал между импульсами должен быть несколько больше абсолютного рефрактерного периода. В двигательных нервных клетках теплокровных животных рефрактерный период составляет около 0,4 мсек, и потенциальный максимальный ритм должен бы быть равным 2500/сек, но на самом деле он равен около 1000/сек. Следует подчеркнуть, что эта частота значительно превышает частоту импульсов, проходящих по этим волокнам в физиологических условиях. Последняя составляет около 100/сек.
Дело в том, что обычно в естественных условиях ткань работает с так называемым оптимальным ритмом. Для пропускания импульсов с таким ритмом не требуется большой силы раздражения. Исследования показали, что частота раздражения и реобаза тока, способного вызывать нервные импульсы с такой частотой, находятся в своеобразной зависимости: реобаза вначале падает по мере роста частоты импульсов, затем снова нарастает. Оптимум находится у нервов в пределах от 75 до 150 имп/сек, для мышц — 20-50 имп/сек. Такой ритм, в отличие от других, очень стойко и длительно может воспроизводиться возбудимыми образованиями.
Таким образом, мы можем теперь назвать все основные параметры возбудимости тканей, характеризующие ее свойства: РЕОБАЗА, ПОЛЕЗНОЕ ВРЕМЯ (ХРОНАКСИЯ), КРИТИЧЕСКИЙ НАКЛОН, ЛАБИЛЬНОСТЬ. Все они, кроме последнего, находятся с возбудимостью в обратно пропорциональных отношениях.
Понятие о «парабиозе«. Лабильность — величина непостоянная. Она может меняться в зависимости от состояния нерва или мышцы, в зависимости от силы и длительности падающих на них раздражений, от степени утомления и т.п. Впервые изменение лабильности нерва при действии на него сначала химическими, а затем и электрическими раздражителями, изучил Н.Е.Введенский. Он обнаружил закономерное снижение лабильности альтерированного химическим агентом (аммиаком) участка нерва, назвал это явление «парабиозом» и изучил его закономерности. Парабиоз — это обратимое состояние, которое, однако, при углублении действия вызывающего его агента может перейти в необратимое.
Лабильность — величина непостоянная. Она может меняться в зависимости от состояния нерва или мышцы, в зависимости от силы и длительности падающих на них раздражений, от степени утомления и т.п. Впервые изменение лабильности нерва при действии на него сначала химическими, а затем и электрическими раздражителями, изучил Н.Е.Введенский. Он обнаружил закономерное снижение лабильности альтерированного химическим агентом (аммиаком) участка нерва, назвал это явление «парабиозом» и изучил его закономерности. Парабиоз — это обратимое состояние, которое, однако, при углублении действия вызывающего его агента может перейти в необратимое.
Введенский рассматривал парабиоз как особое состояние стойкого не колеблющегося возбуждения, как бы застывшего в одном участке нервного волокна. Действительно, парабиотический участок заряжен отрицательно. Такое явление Введенский считал прообразом перехода возбуждения в торможение в нервных центрах. По его мнению, парабиоз является результатом перевозбуждения нервной клетки слишком сильным или слишком частым раздражением.
Развитие парабиоза протекает в три стадии: уравнительную, парадоксальную и тормозную. Вначале за счет снижения аккомодации отдельные импульсы тока малой частоту при условии их достаточной силы дают уже не 1 импульс, а 2,3 или даже 4. Одновременно порог возбудимости растет, а максимальный ритм возбуждения прогрессивно снижается. В результате на импульсы как малой, так и большой частоты нерв начинает отвечать одной и той же частотой разрядов, наиболее близкой к оптимальному для этого нерва ритму. Это и есть уравнительная фаза парабиоза. На следующем этапе развития процесса в области пороговых интенсивностей раздражения еще сохраняется воспроизведение ритма, близкого к оптимальному, а на частые импульсы ткань или вообще не отвечает, или отвечает очень редкими волнами возбуждения. Это — парадоксальная фаза.
Затем способность волокна к ритмической волновой деятельности падает, падает и амплитуда ПД, увеличивается его длительность. , Любое внешнее воздействие подкрепляет состояние торможения нервного волокна и одновременно затормаживается само. Это — последняя, тормозная фаза парабиоза.
, Любое внешнее воздействие подкрепляет состояние торможения нервного волокна и одновременно затормаживается само. Это — последняя, тормозная фаза парабиоза.
В настоящее время описанный феномен объясняется с позиций мембранной теории нарушением механизма повышения натриевой проницаемости и появлением затяжной натриевой инактивации. В результате этого Na-каналы остаются закрытыми, он накапливается в клетке и наружная поверхность мембраны длительное время сохраняет отрицательный заряд. Это препятствует новому раздражению за счет удлинения рефрактерного периода. При набегании на участок парабиоза часто следующих друг за другом ПД, инактивация натриевой проницаемости, вызванная альтерирующим агентом, суммируется с инактивацией, сопровождающей нервный импульс. В результате возбудимость снижается настолько, что проведение следующего импульса полностью блокируется.
Обмен веществ и энергии при возбуждении. При возникновении и проведении возбуждения в нервных клетках и мышечных волокнах происходит усиление обмена веществ. Это проявляется как в ряде биохимических изменений, происходящих в мембране и протоплазме клеток, так и в усилении их теплопродукции. Установлено, что при возбуждении происходят: усиление распада в клетках богатых энергией соединений — АТФ и креатинфосфата (КФ), усиление процессов распада и синтеза углеводов, белков и липидов, усиление окислительных процессов, приводящих в сочетании с гликолизом к резинтезу АТФ и КФ, синтез и разрушение ацетилхолина и норадреналина, других медиаторов, усиление синтеза РНК и белков. Все эти процессы более всего выражены в период восстановления состояния мембраны после ПД.
Это проявляется как в ряде биохимических изменений, происходящих в мембране и протоплазме клеток, так и в усилении их теплопродукции. Установлено, что при возбуждении происходят: усиление распада в клетках богатых энергией соединений — АТФ и креатинфосфата (КФ), усиление процессов распада и синтеза углеводов, белков и липидов, усиление окислительных процессов, приводящих в сочетании с гликолизом к резинтезу АТФ и КФ, синтез и разрушение ацетилхолина и норадреналина, других медиаторов, усиление синтеза РНК и белков. Все эти процессы более всего выражены в период восстановления состояния мембраны после ПД.
В нервах и мышцах каждая волна возбуждения сопровождается выделением двух порций тепла, из которых первая называется начальным, а вторая — запаздывающим теплом. Начальное теплообразование происходит в момент возбуждения и составляет незначительную часть всей теплопродукции (2-10%) при возбуждении. Предполагают, что это тепло связано с теми физико химическими процессами, которые развиваются в момент генерации ПД. Запаздывающее теплообразование происходит в течение более продолжительного времени, и длится многие минуты. Оно связано с теми химическими процессами, которые происходят в ткани вслед за волной возбуждения, и , по образному выражению Ухтомского, составляют «метаболический хвост кометы возбуждения».
Запаздывающее теплообразование происходит в течение более продолжительного времени, и длится многие минуты. Оно связано с теми химическими процессами, которые происходят в ткани вслед за волной возбуждения, и , по образному выражению Ухтомского, составляют «метаболический хвост кометы возбуждения».
Проведение возбуждения. Классификация нервных волокон. Как только в какой-либо точке нервного или мышечного волокна возникает ПД и этот участок приобретает отрицательный заряд, между возбужденными и соседними покоящимися участками волокна возникает электрический ток. В данном случае возбужденный участок мембраны действует на соседние участки как катод постоянного тока, вызывая их деполяризацию и генерируя локальный ответ. Если величина локального ответа превысит Ек мембраны, возникает ПД. В результате наружная поверхность мембраны заряжается отрицательно на новом участке. Таким способом волна возбуждения распространяется вдоль всего волокна со скоростью около 0,5-3 м/сек.
Законы проведения возбуждения по нервам.
1. Закон физиологической непрерывности. Перерезка, перевязка, а также любое другое воздействие, нарушающее целость мембраны (физиологическую, а не только анатомическую), создают непроводимость. То же возникает при тепловых и химических воздействиях.
2. Закон двустороннего проведения. При нанесении раздражения на нервное волокно возбуждение распространяется по нему в обеих направлениях ( по поверхности мембраны — во все стороны) с одинаковой скоростью. Это доказывается опытом Бабухина и подобными ему.
3. Закон изолированного проведения. В нерве импульсы распространяются по каждому волокну изолированно, т.е. не переходят с одного волокна на другое. Это очень важно, так как обеспечивает точную адресовку импульса. Связано это с тем, что электрическое сопротивление миэлиновых и швановской оболочек, а также межклеточной жидкости значительно больше, чем сопротивление мембраны нервных волокон.
Механизмы и скорость проведения возбуждения в безмякотных и мякотных нервных волокнах различны. В безмякотных возбуждение распространяется непрерывно вдоль всей мембраны от одного возбужденного участка к другому, расположенному рядом, так, как мы уже обсуждали.
В миэлиновых волокнах возбуждение распространяется только скачкообразно, перепрыгивая через участки, покрытые миэлиновой оболочкой (сальтаторно). Потенциалы действия в этих волокнах возникают только в перехватах Ранвье. В состоянии покоя наружная поверхность возбудимой мембраны всех перехватов Ранвье заряжена положительно. В момент возбуждения поверхность первого перехвата становится отрицательно заряженной по отношению к соседнему второму перехвату. Это приводит к возникновению местного (локального) электротока, который идет через окружающую волокно межклеточную жидкость, мембрану и аксоплазму от перехвата 2 к 1. Выходящий через перехват 2 ток возбуждает его, вызывая перезарядку мембраны. Теперь этот участок может возбудить следующий и т. д.
д.
Перепрыгивание ПД через межперехватный участок возможно потому, что амплитуда ПД в 5-6 раз больше порога, необходимого для возбуждения не только следующего, но и 3-5 перехватов. Поэтому микроповреждения волокна в межперехватных участках или не одном перехвате не прекращают работы нервного волокна до тех пор, пока регенеративный явления не захватят 3 и более лежащих рядом швановские клетки.
Время, необходимое для передачи возбуждения от одного перехвата к другому, одинаково у волокон различного диаметра, и составляют 0,07 мсек. Однако поскольку длина межперехватных участков различна и пропорциональна диаметру волокна, в миэлинизированных нервах скорость проведения нервных импульсов прямо пропорциональная их диаметру.
Классификация нервных волокон. Электрический ответ целого нерва является алгебраической суммой ПД отдельных его нервных волокон. Поэтому, с одной стороны, амплитуда электрических импульсов целого нерва зависит от силы раздражителя (с ее ростом вовлекаются все новые волокна), а во-вторых, суммарный потенциал действия нерва может быть расчленен на несколько отдельных колебаний, причиной чего является неодинаковая скорость проведения импульсов по разным волокнам, составляющим целый нерв.
В настоящее время нервные волокна по скорости проведения возбуждения, длительности различных фаз ПД и строении принято разделять на три основных типа.
Волокна типа А делятся на подгруппы (альфа, бета, гамма, дельта). Они покрыты миэлиновой оболочкой. Скорость проведения у них самая большая — 70-120 м/сек. Это — двигательные волокна, от моторных нейронов спинного мозга. Остальные волокна типа А — чувствительные.
Волокна типа В — миэлиновые, преимущественно преганглионарные. Скорость проведения — 3-18 м/сек.
Волокна типа С — безмякотные, очень малого диаметра (2 мк). Скорость проведения не больше 3 м/сек. Это постганглионарные волокна симпатической нервной системы чаще всего.
Тема 5. Действие постоянного тока на живые ткани. Хронаксиметрия . Полный курс за 3 дня. Нормальная физиология
Постоянный ток в настоящее время находит все более широкое применение в клинической практике, как для диагностики поражений нервов и мышц (например, хронаксиметрия – метод определения возбудимости периферических нервов и скелетных мышц), так и для физиотерапии ряда заболеваний (например, использование постоянного тока для введения лекарственных веществ – метод электрофореза – или с целью повышения эластичности послеоперационного рубца).
Постоянный ток – это ток, постоянный по силе и направлению. Он меняет свою величину только дважды – в момент замыкания цепи (при этом амплитуда резко возрастает с нуля до определенного значения) и в момент размыкания цепи (при этом амплитуда резко снижается с определенной величины до нуля). Таким образом, постоянный ток, в отличие от переменного, будет действовать на живые ткани только в момент замыкания и размыкания цепи. После замыкания цепи и ответной реакции в тканях начинается адаптация к действию постоянного тока.
Известно, что в момент замыкания цепи постоянного тока возбуждение возникает под катодом, а при размыкании – под анодом (Полярный закон Пфлюгера). В 1859 г. Пфлюгер провел следующий опыт. Умерщвляя участок нерва под одним из электродов и устанавливая на неповрежденный участок другой электрод, он обнаружил, что при соприкосновении с неповрежденным участком катода возбуждение возникает только при замыкании цепи постоянного тока, а если катод установить на поврежденный участок ткани, а анод на неповрежденный, то возбуждение возникает только при размыкании цепи.
В момент замыкания цепи в области приложения к поверхности ткани положительно заряженного анода увеличивается положительный потенциал на наружной поверхности клеточной мембраны, т. е. происходит ее гиперполяризация, при этом увеличивается мембранный потенциал, поэтому при замыкании цепи постоянного тока возбуждение над анодом не возникает. Это явление не сопровождается изменением ионной проницаемости клеточных мембран и получило название пассивной гиперполяризации.
В момент замыкания цепи в области приложения отрицательно заряженного электрода – катода – положительный заряд на наружной поверхности клеточной мембраны снижается. Возникают пассивная деполяризация и снижение величины мембранного потенциала. В момент замыкания цепи повышается проницаемость мембраны для ионов натрия, что увеличивает явление деполяризации, что в свою очередь способствует еще большему увеличению натриевой проницаемости.
Возникают пассивная деполяризация и снижение величины мембранного потенциала. В момент замыкания цепи повышается проницаемость мембраны для ионов натрия, что увеличивает явление деполяризации, что в свою очередь способствует еще большему увеличению натриевой проницаемости.
Прохождение постоянного электрического тока через живую ткань сопровождается изменением ее физических и химических свойств. Для обозначения этих изменений введен термин «электротон». Изменения, которые происходят над катодом, получили название «катэлектротон», под анодом – «анэлектротон». Изменения, возникающие на расстоянии 1 см от электронов, называются «периэлектротон», они противоположны изменениям, возникающим на катоде и на аноде.
В момент замыкании цепи происходят определенные сдвиги физиологических и физико-химических свойств.
Под катодом

Под анодом понижаются возбудимость и проводимость ткани, повышается активность холинэстеразы, уменьшается содержание ацетилхолина, накапливается витамин В2, выделяется углекислый газ.
При продолжительном действии постоянного электрического тока под катодом увеличивается критический уровень деполяризации, т. е. возрастает порог раздражения. Наряду с этим происходит снижение амплитуды потенциала действия, так как длительное повышение натриевой проницаемости над катодом в момент замыкания цепи приводит к ее аккомодации. Накопление под катодом ацетилхолина также способствует понижению возбудимости за счет развития стойкой деполяризации. Это явление – повышение возбудимости над катодом, которое затем сменяется ее снижением, получило название катодической депрессии и было изучено учеником Н. Е. Введенского Б. Ф. Вериго.
В зависимости от расположения электродов различают восходящее и нисходящее направление тока. При восходящем направлении ближе к мышце располагается анод, а при нисходящем – катод.
Ответная реакция ткани зависит не только от направления постоянного тока, но и от его силы. Различают слабый (пороговый), средний и сильный постоянный ток.
Слабый ток вызывает ответную реакцию в мышце или при восходящем или при нисходящем направлении только в момент замыкания цепи. В момент размыкания цепи ответной реакции не возникает, так как под анодом развивается только местное возбуждение, которое не проводится к мышце.
Средний ток при восходящем и при нисходящем направлении вызывает ответную реакцию как при замыкании, так и при размыкании цепи. В момент замыкания цепи под катодом, а в момент размыкания – под анодом возникает импульсное возбуждение, которое и вызывает сокращение мышцы.
Сильный ток при восходящем направлении вызывает ответную реакцию при размыкании цепи, а при нисходящем – только в момент ее замыкания, когда возбуждение возникает под электродом, расположенным ближе к мышце.
В момент замыкания цепи сильного постоянного тока восходящего направления под катодом возникает распространяющееся возбуждение, но в момент действия сильного постоянного тока под анодом резко понижаются возбудимость и проводимость, что блокирует проведение возбуждения от катода через область анода.
Аналогичный блок проведения возбуждения создается под анодом при размыкании цепи постоянного тока нисходящего направления за счет возникновения катодической депрессии.
Хронаксиметрия – один из методов диагностики функционального состояния нервов и мышц с помощью постоянного электрического тока. Для характеристики возбудимости ткани необходимо определить минимальный порог раздражения (реобазу) и минимальное время, в течение которого ток, по силе или по напряжению равный удвоенной реобазе, должен возбудить ткань (хронаксия).
Хронаксия – это величина, характеризующая скорость возникновения возбуждения в ткани. Чем быстрее возбуждается ткань, тем короче ее хронаксия. Хронаксия измеряется в тысячных долях секунды, реобаза – в вольтах или миллиамперах.
Лабильность и хронаксия тесно связаны между собой, так как быстро протекающий процесс возбуждения характеризуется быстрым возникновением, и, наоборот, медленное протекание процесса возбуждения сочетается с длительным его возникновением. Таким образом, измерение хронаксии можно использовать для характеристики лабильности тех или иных образований. Хронаксия и лабильность ткани находятся в обратно пропорциональной зависимости. Чем больше хронаксия, тем меньше лабильность ткани, и наоборот – при низкой хронаксии лабильность ткани высокая.
Таким образом, измерение хронаксии можно использовать для характеристики лабильности тех или иных образований. Хронаксия и лабильность ткани находятся в обратно пропорциональной зависимости. Чем больше хронаксия, тем меньше лабильность ткани, и наоборот – при низкой хронаксии лабильность ткани высокая.
Для определения хронаксии пользуются прибором хронаксометром. Он позволяет дозировать время действия тока на ткань и его силу. В хронаксометре имеются два электрода, отличающиеся друг от друга по размерам: анод – большой электрод, катод – малый электрод. В связи с этим густота электрических линий у анода незначительна и раздражающий эффект практически отсутствует, поэтому большой электрод называется индифферентным. Густота электрических линий на катоде примерно в 100 раз больше, чем на аноде, и он обладает выраженным раздражающим действием. Этот электрод называется дифферентным, или активным.
При исследовании проводят определение хронаксии эфферентных (моторную хронаксию) и афферентных (сенсорную хронаксию – зрительную, слуховую) систем.
При исследовании мышц раздражение наносится на их двигательную точку – проекцию на коже места входа нервного ствола в данную мышцу. Для обнаружения этих точек используют системы их топографии. Передача возбуждения с одного нейрона на другой, а также с нейрона на мышцу возможна только при близких величинах их хронаксии. Это явление получило название изохромизма. Если хронаксии мышцы и нерва отличаются друг от друга более чем в два раза, то передача возбуждения невозможна, что получило название гетерохромизма.
Хронаксия тканей – непостоянная величина и зависит от многих факторов.
Хронаксия периферических нервов зависит от состояния центров в спинном мозге и в вышележащих отделах головного мозга. Хронаксия изолированного нервного волокна значительно увеличивается. Создание доминанты в коре головного мозга, травма головного мозга увеличивают хронаксию мышц и нервов конечностей. Физическая активность вначале укорачивает хронаксию, а при развитии утомления удлиняет ее. При действии холода и развитии утомления хронаксия удлиняется и наступает явление гетерохромизма между мышцей и нервом. Во время сна хронаксия мышц-разгибателей удлиняется. При наличии болевого синдрома в мышечно-суставном аппарате хронаксия укорачивается. При уменьшении болей она постепенно нормализуется.
Хронаксия изолированного нервного волокна значительно увеличивается. Создание доминанты в коре головного мозга, травма головного мозга увеличивают хронаксию мышц и нервов конечностей. Физическая активность вначале укорачивает хронаксию, а при развитии утомления удлиняет ее. При действии холода и развитии утомления хронаксия удлиняется и наступает явление гетерохромизма между мышцей и нервом. Во время сна хронаксия мышц-разгибателей удлиняется. При наличии болевого синдрома в мышечно-суставном аппарате хронаксия укорачивается. При уменьшении болей она постепенно нормализуется.
Данный текст является ознакомительным фрагментом.
Законы действия постоянного тока на возбудимые ткани
Похожие презентации:
Механизм развития потенциала действия. Строение и работа ионных каналов. Распространение потенциала действия вдоль нервного волокна
Нормальная Физиология для ММА. Физиология возбудимых тканей
Физиология возбудимых тканей
Законы раздражения возбудимых тканей и законы возбуждения
Электротерапия: постоянный ток, импульсные токи
Основы электрофизиологии. Общая характеристика возбудимых тканей. Потенциал покоя. Потенциал действия
Потенциал действия
Физиология возбудимых тканей
Законы раздражения возбудимых тканей. Законы проведения возбуждения по нервам
Физиология. Физиология возбудимых тканей. (Лекция 1)
Введение в курс физиологии. Основные принципы формирования и регуляции физиологических функций. Природа возбуждения и торможения
1. Лекция №3 Законы действия постоянного тока на возбудимые ткани
2Отличия локального
ответа от потенциала
действия:
Локальный ответ
1.
2.
3.
4.
5.
Eк
Eo
Возникает на
подпороговые
раздражители.
Не распространяется.
Не подчиняется
правилу
“Все или
ничего”.
Способен к суммации.
Возбудимость в
период локального
ответа повышена.

Если локальный ответ
достигнет уровня
критической
деполяризации, он
перерастает в
потенциал действия.
3
Основные электрофизиологические феномены в
нервном волокне
Аэт — анаэлектротон, КУД — критический уровень деполяризации, Кэт —
катэлектротон, ЛО — локальный (подпороговый активный) ответ, МПП —
мембранный потенциал покоя, ПД — потенциал действия, СП (отр и пол)
— следовые потенциалы отрицательный и положительный (временные
соотношения пика ПД и СП не выдержаны; отрицательный СП и
особенно положительный СП значительно длительнее).
4. 4 Отличие локального ответа от электротонического потенциала
мВ+40
ПД
0
ЛО
КУД
ЭП
ПП
-80
мА
мс
2
4
6
5. 5 Локальные ответы и закон силы
Сила раздражителя в вольтах0,5
1
1,0
1,5
2
3
2,0
4
0
Порог
Ек
Е0
— 90
1
2
3
4
7. 7 ЦИКЛ ХОДЖКИНА-ХАКСЛИ
7ЦИКЛ
Раздражитель
ХОДЖКИНА-ХАКСЛИ
деполяризация мембраны
возрастание
входящего
Na+ — тока
повышение Na+
проницаемости
8
Изменение возбудимости клетки в
разные фазы ПД
а — мембранный
потенциал (исходная
возбудимость),
б — локальный ответ
(повышенная
возбудимость),
в — потенциал действия
(абсолютная и
относительная
рефрактерность),
г — следовая
деполяризация
(супернормальная
возбудимость),
д — следовая
гиперполяризация
(субнормальная
возбудимость)
СОСТОЯНИЕ НАТРИЕВЫХ КАНАЛОВ
А
Na+
А
ИнА
ИнА
СОСТОЯНИЕ
ПОТЕНЦИАЛА
ПОКОЯ
СОСТОЯНИЕ
ДЕПОЛЯРИЗАЦИИ
МЕМБРАНЫ
СПАЙК И
РЕПОЛЯРИЗАЦИЯ
МЕМБРАНЫ
10.
 10 Полярный закон ПфлюгераВозбуждение возникает в момент
10 Полярный закон ПфлюгераВозбуждение возникает в моментзамыкания цепи под катодом, а в
момент размыкания цепи под
анодом.
11. 11 Полярный закон действия тока – возбуждение возникает под катодом при замыкании и под анодом при размыкании тока
Цепь событий, развивающихся под катодомраздражающего тока:
пассивная деполяризация мембраны
повышение натриевой проницаемости
усиление потока Na++ внутрь волокна
активная деполяризация мембраны
локальный ответ
достижение критического уровня (Ек )
регенеративная деполяризация
потенциал действия (ПД).
12. 12 Пассивные изменения
КАТОДАНОД
«-»
«+»
———————
——-+++——-++++
——
13. 13 Пассивные изменения
14. 14 Закон физиологического электротона
В момент замыкания цепи возбудимость ипроводимость под катодом
увеличиваются – катэлектротон;
а под анодом – уменьшаются –
анэлектротон;
При размыкании цепи возбудимость под
катодом уменьшается – обратный
катэлектротон;
а под анодом – увеличивается – обратный
анэлектротон.
15. 15 Катэлектротон и анэлектротон
16. 16 Катодическая депрессия Вериго (1889 г.)
При длительно
действующей
деполяризации током
мембраны, развиваются
процессы повышающие
критический уровень
деполяризации. Это связано
с развитием инактивации
натриевых каналов и
активации калиевых.
Одновременно с
увеличением порога,
происходит снижение
амплитуды ПД и крутизны
его нарастания.
17
Изменения возбудимости при
длительном действии катода
Катодическая депрессия
Вериго
Катодзамыкательное
возбуждение
Ек-2
Ек-1
Ео
замыкание
размыкание
18
Изменения возбудимости при
длительном действии анода
Ек-1
Ек-2
Ео
замыкание
размыкание
19. 19 Возникновение анод-размыкательного возбуждения
20. 20 Аккомодация
• Аккомодация (от лат. Accommodatioприспособление, приноровление)-общеесвойство возбудимых тканей.
• Аккомодация- повышение порога
возбуждения к медленно нарастающему или
постоянно действующему раздражителю.

• Закон крутизны нарастания раздражителя
(Законы раздражения): Чем выше крутизна
нарастания раздражителя во времени, тем
больше до известного предела величина
функционального ответа.
21. 21 Аккомодация
В основе аккомодациилежит развитие
постепенной
инактивация
натриевых каналов и
повышение калиевой
проводимости,
возникающие во время
медленно нарастающей
деполяризации
мембраны.
22. 22 Парабиоз
Местное нераспростроняющееся иуглубляющееся во времени возбуждение.
Обнаружен Введенским при исследовании
способности
нерва
проводить
высокочастотные разряды импульсов после
воздействия на нерв различных химических
агентов.
В
дальнейшем
было
показано,
что
состояние
парабиоза
можно
вызвать
действием раздражителя любой природы.
23. 23 Парабиоз и его фазы
Парабиоз и его фазы23
• Уравнительная
• Парадоксальная
• Тормозная
нерв
Nh5
сила раздражения (Гц)
• Введенский впервые обратил внимание на то, что
способность ткани воспроизводить задаваемый ритм
связана с ее функциональным состоянием –
лабильностью.

• Явление парабиоза широко распространено в
природе
–
зимняя
спячка
(анабиоз),
как
хладнокровных, так и теплокровных (медведи).
• Использование наркотических веществ позволяет
проводить многочасовые операции, а анестетики –
блокируют болевые ощущения.
• Открытие парабиоза, а затем пессимального
торможения, позволило Введенскому выдвинуть
положение о тормозных явлениях в нервах, как о
стойком нераспростроняющемся возбуждении.
27. Проведение возбуждения по нервному волокну
Законы проведениявозбуждения по нерву
28. 28 Образование миелинового волокна
29 Миелиновое нервное волокномиелин
перехваты Ранвье
30 Скорость передачи сигнала по
нервным волокнам
32. В основе распространения возбуждения — возникновение местных токов между деполяризованным и покоящимся участками. Между различно заряжен
В основе распространения возбуждения — возникновение местных токовмежду деполяризованным и покоящимся участками.

Между различно заряженными участками мембраны возникает локальный
ионный ток, который деполяризует мембрану до критического уровня. Причем
подпороговый деполяризующий мембрану ток идет по аксоплазме , то есть — изнутри.
В результате этой подпороговой деполяризации открываются Nа-каналы и
возрастает входящий Nа ток.
Отношение величины ПД к пороговому току называется фактором
надежности:
Амплитуда ПД, мВ
Величина Екр, мВ
Плотность Nа+-каналов в перехватах Ранвье: 10 000/мм2, что в 200 раз
больше, чем в нервном волокне гигатского аксона кальмара.
Входящий Nа+-ток, пронизывающий невозбужденную мембрану в
непосредственной близости от ее возбужденного участка в 5-6 раз выше
порогового тока. То есть фактор надежности настолько высок, что позволяет
перескакивать ПД через несколько перехватов Ранвье, при их блокаде
анестетиками.
33. 33 Законы проведения возбуждения по нерву
Законы проведениявозбуждения по нерву
33
1.
 Закон физиологической
Закон физиологическойнепрерывности
2. Закон двустороннего проведения
3. Закон изолированного проведения
4. Закон бездекрементного
(незатухающего) проведения
возбуждения
5. Закон относительной неутомляемости
нерва (открыт Введенским)
34. 34 Двустороннее проведение возбуждения экспериментально доказано:
Двустороннее проведениевозбуждения экспериментально
доказано:
34
Бабухиным А.И. (1877) на
электрическом органе нильского
сома.
Кюне В. (1886) на икроножной
мышце лягушки.
35. 35 Опыт Бабухина А.И.
12
3
4
5
6
36. 36 Опыт Бабухина А.И.
12
3
разрез
4
разрез
5
6
раздражение
37. 37 Опыт Кюне В.
Опыт Кюне В.37
разрез
разрез
38. 38 Классификация нервных волокон
• Волокна типа А (ά, β, δ) – мякотныетолстые моторные волокна, скорость
проведения возбуждения до 120
м/сек.
• Волокна типа В –тонкие мякотные
волокна, чаще чувствительные,
скорость проведения 3-18 м/сек.

• Волокна типа С – безмякотные,
вегетативные, скорость проведения
не больше 3 мсек.
39
Тип
Типы нервных волокон, их
свойства и функциональное
назначение
Диаметр
(мкм)
Миелинизация
Скорость
проведения
(м/с)
Функциональное назначение
А
12–20
сильная
70–120
Двигательные волокна соматической НС;
чувствительные волокна
проприорецепторов
А
5–12
сильная
30–70
Чувствительные волокна кожных
рецепторов
А
3–16
сильная
15–30
Чувствительные волокна
проприорецепторов
А
2–5
сильная
12–30
Чувствительные волокна терморецепторов,
ноцицепторов
В
1–3
слабая
3–15
Преганглионарные волокна симпатической
НС
С
0,3–1,3
отсутствует
0,5–2,3
Постганглионарные волокна
симпатической НС; чувствительные
волокна терморецепторов, ноцицепторов,
некоторых механорецепторов
English Русский Правила
Нейробиологические механизмы транскраниальной стимуляции постоянным током при психических расстройствах; Нейрофизиологические, химические и анатомические аспекты
Введение
Транскраниальная стимуляция постоянным током (tDCS) — это неинвазивный метод, который модулирует активность нейронов в головном мозге путем подачи низкоамплитудного (обычно не более 2 мА) импульса в течение короткого периода ( не более 30 мин) между электродами (анод и катод). По крайней мере, один из электродов помещается на скальп, через который электронные токи проникают через череп в мозг и облегчают или подавляют спонтанную нейронную активность вблизи электродов (Yokoi et al., 2018; рисунок 1).
По крайней мере, один из электродов помещается на скальп, через который электронные токи проникают через череп в мозг и облегчают или подавляют спонтанную нейронную активность вблизи электродов (Yokoi et al., 2018; рисунок 1).
Рисунок 1 . Схематическая диаграмма (слева) и экспериментальная установка (справа) для tDCS. (слева) Анодный и катодный электроды располагаются над левой дорсолатеральной префронтальной корой и над правой надглазничной областью соответственно. Направление тока — от анода к катоду. (справа) Администратор управляет стимулятором (а). Анодный (б) и катодный (в) электроды размером 35 см 2 накладывают на F3 и правую надглазничную область соответственно. Головной ремень (d) используется по мере необходимости для повышения воспроизводимости.
Сообщалось об эффективности tDCS при лечении большого депрессивного расстройства (БДР) (Yokoi et al., 2018).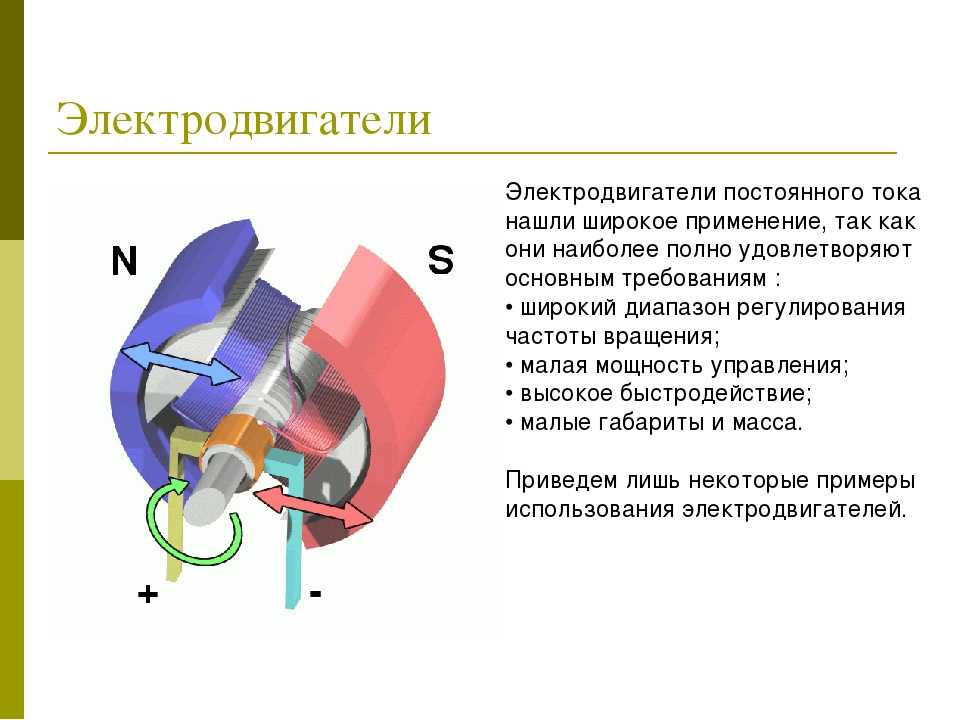 Так, метаанализ показал умеренное влияние tDCS на симптомы депрессии у пациентов с острой депрессией (Hedges’g = 0,37) (Shiozawa et al., 2014). Кроме того, была серия отчетов, показывающих способность tDCS улучшать положительные/отрицательные симптомы шизофрении (Kim et al., 2019). Например, было обнаружено, что галлюцинации (положительные симптомы) (Hedges’g = 0,86) и негативные симптомы (0,41) улучшаются при многократных сеансах tDCS на лобных или лобно-височных долях (см. монтаж в таблице 1) два раза в день в течение 5 дней. (Ким и др., 2019 г.). Кроме того, метаанализ когнитивной функции у пациентов с шизофренией указывает на способность многосеансовой tDCS на префронтальной коре (см. монтаж в таблице 1) улучшать рабочую память (Hedges’g = 0,49), важную когнитивную область (Narita et al. др., 2020; табл. 1).
Так, метаанализ показал умеренное влияние tDCS на симптомы депрессии у пациентов с острой депрессией (Hedges’g = 0,37) (Shiozawa et al., 2014). Кроме того, была серия отчетов, показывающих способность tDCS улучшать положительные/отрицательные симптомы шизофрении (Kim et al., 2019). Например, было обнаружено, что галлюцинации (положительные симптомы) (Hedges’g = 0,86) и негативные симптомы (0,41) улучшаются при многократных сеансах tDCS на лобных или лобно-височных долях (см. монтаж в таблице 1) два раза в день в течение 5 дней. (Ким и др., 2019 г.). Кроме того, метаанализ когнитивной функции у пациентов с шизофренией указывает на способность многосеансовой tDCS на префронтальной коре (см. монтаж в таблице 1) улучшать рабочую память (Hedges’g = 0,49), важную когнитивную область (Narita et al. др., 2020; табл. 1).
Таблица 1 . Мета-анализ эффектов tDCS.
Несмотря на накопленные данные об эффективности tDCS при лечении психических расстройств, особенно шизофрении и расстройств настроения, механизм его действия до конца не выяснен (Stagg and Nitsche, 2011). Таким образом, текущий обзор был направлен на то, чтобы предоставить обзор действия tDCS, особенно анодной стимуляции, на нейротрансмиссию и нейронные сети в головном мозге, чтобы помочь понять механизмы, лежащие в основе его терапевтических эффектов.
Таким образом, текущий обзор был направлен на то, чтобы предоставить обзор действия tDCS, особенно анодной стимуляции, на нейротрансмиссию и нейронные сети в головном мозге, чтобы помочь понять механизмы, лежащие в основе его терапевтических эффектов.
О влиянии tDCS на психические симптомы в основном сообщалось в исследованиях с использованием анодной стимуляции лобной коры. С другой стороны, место размещения катодного электрода не было однородным, что указывает на то, что анодная стимуляция вызвала интерес к рассмотрению механизма tDCS (Fregni et al., 2020). Поскольку клинические преимущества tDCS были обнаружены при применении нескольких сеансов (Shiozawa et al., 2014; Kim et al., 2019; Narita et al., 2020), акцент был сделан на долгосрочных изменениях нейронных событий, вызванных по tDCS.
Нейрофизиологическое понимание tDCS
Анодная стимуляция tDCS (1–2 мА) сама по себе недостаточно сильна, чтобы деполяризовать мембранный потенциал нейронов до порога срабатывания, и только увеличивает скорость спонтанного возгорания и их возбудимость (Nitsche и Paulus, 2000; Philip et al.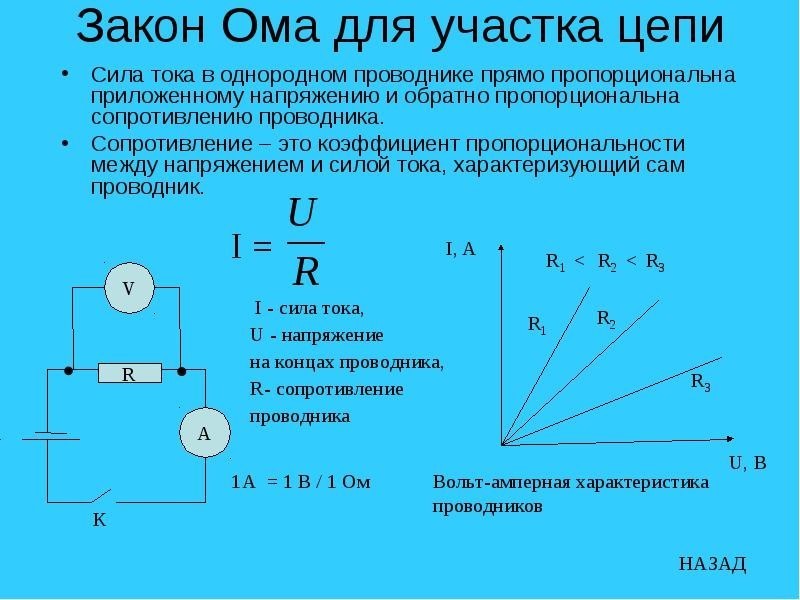 , 2017; рисунок 2). И наоборот, считается, что катодная стимуляция углубляет потенциал покоящейся мембраны, затрудняя деполяризацию нейронов, что снижает скорость спонтанного возгорания и возбудимость нейронов (Nitsche and Paulus, 2000; Philip et al., 2017; рисунок 2). Важно отметить, что эти эффекты tDCS зависят от интенсивности и продолжительности стимуляции (Nitsche and Paulus, 2000) и радиального электрического поля (Seo and Jun, 2019).).
, 2017; рисунок 2). И наоборот, считается, что катодная стимуляция углубляет потенциал покоящейся мембраны, затрудняя деполяризацию нейронов, что снижает скорость спонтанного возгорания и возбудимость нейронов (Nitsche and Paulus, 2000; Philip et al., 2017; рисунок 2). Важно отметить, что эти эффекты tDCS зависят от интенсивности и продолжительности стимуляции (Nitsche and Paulus, 2000) и радиального электрического поля (Seo and Jun, 2019).).
Рисунок 2 . Схематическая диаграмма изменения потенциала нейронов с помощью tDCS. Анодная стимуляция tDCS (1-2 мА) недостаточно сильна, чтобы деполяризовать мембранный потенциал нейронов до порога срабатывания. И наоборот, считается, что катодная стимуляция углубляет мембранный потенциал покоя и затрудняет деполяризацию нейронов.
Электрофизиологическое понимание tDCS может быть облегчено моделью, зависящей от стимуляции (Fertonani et al., 2011). В этой модели считается, что анодная стимуляция способствует деполяризации нейронов, а катодная вызывает гиперполяризацию для ее подавления. Более того, электрическая стимуляция воздействует на несколько нейронов и увеличивает их мембранный потенциал, вызывая деполяризацию. Эти события вблизи нервных мембран были предложены для объяснения способности tDCS улучшать функции мозга (Silvanto et al., 2008).
Более того, электрическая стимуляция воздействует на несколько нейронов и увеличивает их мембранный потенциал, вызывая деполяризацию. Эти события вблизи нервных мембран были предложены для объяснения способности tDCS улучшать функции мозга (Silvanto et al., 2008).
Считается, что повышенная возбудимость местных нейронов при анодной стимуляции увеличивает кровоток вокруг места стимуляции и вызывает последующие метаболические изменения. В частности, изменения кровотока через tDCS в префронтальной коре были измерены с помощью функциональной ближней инфракрасной спектроскопии (fNIRS) (Merzagora et al., 2010). В этом исследовании увеличение концентрации оксигенированного гемоглобина под анодным электродом было значительно больше, чем под катодом. Считается, что это отражает способность анодной стимуляции вызывать метаболические изменения среди нейронов (Merzagora et al., 2010).
Биохимические эффекты tDCS
Сообщалось об изменениях нейротрансмиссии при анодной стимуляции в связи с метаболическими изменениями в головном мозге. Здесь мы рассматриваем накопленные данные об эффекте стимуляции моторной коры у пациентов с хронической болью и тех, кто проходит постинсультную двигательную реабилитацию (Medeiros et al., 2012; таблицы 2, 3). Например, эффект анодной стимуляции подавляется карбамазепином (ингибитором натриевых каналов) (Liebetanz et al., 2002), что указывает на то, что ингибирование внутриклеточного притока внеклеточного иона натрия подавляет вызванную анодом деполяризацию нейронов и последующие возбуждения.
Здесь мы рассматриваем накопленные данные об эффекте стимуляции моторной коры у пациентов с хронической болью и тех, кто проходит постинсультную двигательную реабилитацию (Medeiros et al., 2012; таблицы 2, 3). Например, эффект анодной стимуляции подавляется карбамазепином (ингибитором натриевых каналов) (Liebetanz et al., 2002), что указывает на то, что ингибирование внутриклеточного притока внеклеточного иона натрия подавляет вызванную анодом деполяризацию нейронов и последующие возбуждения.
Таблица 2 . Влияние сопутствующих препаратов на анодную tDCS на моторную кору здоровых субъектов (адаптировано из Medeiros et al., 2012).
Таблица 3 . Фармакологические действия.
Подтипы рецепторов глутамата, управляющие возбуждающей синаптической передачей, включают рецепторы AMPA (α-амино-3-гидрокси-5-метил-4-изоксазолпропионовая кислота) и NMDA (N-метил-D-аспартат), оба из которых связаны с ион- каналы. Рецептор AMPA участвует во внутриклеточном притоке ионов натрия во время деполяризации нейронов, вызывает транзиторные потенциалы действия и отвечает за большую часть возбуждающих синаптических передач. С другой стороны, рецептор NMDA участвует во внутриклеточном притоке ионов кальция во время деполяризации, вызывает пролонгированные потенциалы действия и опосредует нейронные цепи, управляющие памятью и обучением. Следовательно, воздействия на NMDA-рецепторы, индуцирующие пластичность нейронов, играют доминирующую роль в улучшении симптомов психических расстройств. Соответственно, декстрометорфан (ингибитор рецептора NMDA) подавляет эффект анодной стимуляции (Liebetanz et al., 2002; Nitsche et al., 2003, 2004a), тогда как d-циклосерин (частичный агонист рецептора NMDA) продлевает его (Nitsche et al. ., 2004б). Это согласуется с наблюдениями, что агонисты рецепторов NMDA усиливают возбуждающую синаптическую передачу, в то время как ингибиторы рецепторов NMDA подавляют ее (Liebetanz et al., 2002; Nitsche et al., 2003, 2004a,b). Кроме того, определенную роль может играть ГАМК (гамма-аминомасляная кислота: γ-аминомасляная кислота), нейротрансмиттер, подавляющий синаптические передачи. Таким образом, лоразепам, агонист рецептора ГАМК, задерживает эффект анодной стимуляции (Nitsche et al.
С другой стороны, рецептор NMDA участвует во внутриклеточном притоке ионов кальция во время деполяризации, вызывает пролонгированные потенциалы действия и опосредует нейронные цепи, управляющие памятью и обучением. Следовательно, воздействия на NMDA-рецепторы, индуцирующие пластичность нейронов, играют доминирующую роль в улучшении симптомов психических расстройств. Соответственно, декстрометорфан (ингибитор рецептора NMDA) подавляет эффект анодной стимуляции (Liebetanz et al., 2002; Nitsche et al., 2003, 2004a), тогда как d-циклосерин (частичный агонист рецептора NMDA) продлевает его (Nitsche et al. ., 2004б). Это согласуется с наблюдениями, что агонисты рецепторов NMDA усиливают возбуждающую синаптическую передачу, в то время как ингибиторы рецепторов NMDA подавляют ее (Liebetanz et al., 2002; Nitsche et al., 2003, 2004a,b). Кроме того, определенную роль может играть ГАМК (гамма-аминомасляная кислота: γ-аминомасляная кислота), нейротрансмиттер, подавляющий синаптические передачи. Таким образом, лоразепам, агонист рецептора ГАМК, задерживает эффект анодной стимуляции (Nitsche et al. , 2004c). С другой стороны, сам анодный стимул вызывает локальное снижение концентрации ГАМК в коре (Stagg et al., 2009)., 2011).
, 2004c). С другой стороны, сам анодный стимул вызывает локальное снижение концентрации ГАМК в коре (Stagg et al., 2009)., 2011).
Сообщалось, что моноаминовые нейротрансмиттеры, такие как дофамин, серотонин и ацетилхолин, опосредуют эффект tDCS (Nitsche et al., 2006, 2009; Kuo et al., 2007, 2008; Monte-Silva et al., 2010; Тиругнансамбандам и др., 2011). Например, сульпирид, блокатор дофаминовых рецепторов (Nitsche et al., 2006), подавляет действие анодных раздражителей, а леводопа, предшественник дофамина (Kuo et al., 2008; Monte-Silva et al., 2010), локально усиливает возбуждение определенных синаптических передач (Kuo et al., 2008). Эти данные свидетельствуют о том, что действие tDCS может включать регуляцию передачи дофамина. Кроме того, циталопрам, ингибитор обратного захвата серотонина, усиливает анодную стимуляцию (Nitsche et al., 2009).). Что касается передачи ацетилхолина, ривастигмин, ингибитор холинэстеразы, подавляет эффект tDCS (Kuo et al., 2007). Таким образом, направление влияния на действия анодной стимуляции варьируется в зависимости от моноаминовых нейротрансмиттеров.
Приведенные выше соображения в целом приводят к концепции, согласно которой анодная стимуляция усиливает возбуждающие синаптические передачи за счет стимуляции передачи глутамата и подавления передачи ГАМК в коре. С другой стороны, он модулирует дофаминовую систему, усиливает и подавляет передачу серотонина и ацетилхолина соответственно. Считается, что эти эффекты tDCS на передачу моноаминов связаны с изменением баланса между возбуждающими и тормозными входами в головном мозге (Okun and Lampl, 2008).
Нейроанатомическое понимание эффекта tDCS
Сообщалось о нарушении функциональной связи между областями мозга у пациентов с психическими расстройствами, такими как шизофрения и биполярное расстройство (Yamada et al., 2020). У пациентов с шизофренией исследование с использованием функциональной магнитно-резонансной томографии (фМРТ) в состоянии покоя выявило разделение между медиальной префронтальной корой и дорсолатеральной префронтальной корой (Chai et al., 2011). В другом исследовании были обнаружены изменения динамической функциональной связности в основном в таламусе и мозжечке, а также в лобной, височной, затылочной, веретенообразной, постцентральной, клиновидной, супрамаргинальной и шпорной коре у пациентов с шизофренией или биполярным расстройством. В частности, при шизофрении и биполярном расстройстве ослабляются функциональные связи, включающие постцентральную, лобную и мозжечковую кору, в то время как связи, включающие островковую, височную, лобную, веретенообразную, язычную, затылочную, супрамаргинальную кору, а также таламус и мозжечок, ослабевают. усилены (Du et al., 2017). Кунце и др. систематически применял tDCS к крупномасштабной сетевой модели, состоящей из 74 областей мозга, для исследования функциональной связи динамических состояний. Они обнаружили изменения конкурентных взаимосвязей функциональных сетей с помощью tDCS (Kunze et al., 2016).
В другом исследовании были обнаружены изменения динамической функциональной связности в основном в таламусе и мозжечке, а также в лобной, височной, затылочной, веретенообразной, постцентральной, клиновидной, супрамаргинальной и шпорной коре у пациентов с шизофренией или биполярным расстройством. В частности, при шизофрении и биполярном расстройстве ослабляются функциональные связи, включающие постцентральную, лобную и мозжечковую кору, в то время как связи, включающие островковую, височную, лобную, веретенообразную, язычную, затылочную, супрамаргинальную кору, а также таламус и мозжечок, ослабевают. усилены (Du et al., 2017). Кунце и др. систематически применял tDCS к крупномасштабной сетевой модели, состоящей из 74 областей мозга, для исследования функциональной связи динамических состояний. Они обнаружили изменения конкурентных взаимосвязей функциональных сетей с помощью tDCS (Kunze et al., 2016).
На основании этих результатов механизм действия tDCS на нейронные цепи резюмирован в таблице 4. Анодальный tDCS может усиливать возбуждающую синаптическую передачу путем изменения баланса между активностью глутамата и ГАМК (Clark et al., 2011; Stagg et al. , 2014; Bachtiar et al., 2015; Hunter et al., 2015), что приводит к модификации функциональной связи между областями мозга, включая место стимуляции (Polanía et al., 2011; Stagg et al., 2014; Bachtiar et al. ., 2015; Хантер и др., 2015). Кроме того, эффекты tDCS могут распространяться на головной мозг за счет повышенного/пониженного высвобождения трансмиттеров моноаминов, таких как дофамин, в нейронных цепях, которые не обязательно включают сайт анодной стимуляции (Polanía et al., 2011; Hunter et al. ., 2015; Фонтено и др., 2018). Считается, что эти нейронные события улучшают психиатрические симптомы и когнитивную функцию (Fukai et al., 2019).). Таким образом, анодная стимуляция, вероятно, изменяет уровни активности как определенных областей мозга, так и множественных сетевых систем (Luft et al., 2014).
Анодальный tDCS может усиливать возбуждающую синаптическую передачу путем изменения баланса между активностью глутамата и ГАМК (Clark et al., 2011; Stagg et al. , 2014; Bachtiar et al., 2015; Hunter et al., 2015), что приводит к модификации функциональной связи между областями мозга, включая место стимуляции (Polanía et al., 2011; Stagg et al., 2014; Bachtiar et al. ., 2015; Хантер и др., 2015). Кроме того, эффекты tDCS могут распространяться на головной мозг за счет повышенного/пониженного высвобождения трансмиттеров моноаминов, таких как дофамин, в нейронных цепях, которые не обязательно включают сайт анодной стимуляции (Polanía et al., 2011; Hunter et al. ., 2015; Фонтено и др., 2018). Считается, что эти нейронные события улучшают психиатрические симптомы и когнитивную функцию (Fukai et al., 2019).). Таким образом, анодная стимуляция, вероятно, изменяет уровни активности как определенных областей мозга, так и множественных сетевых систем (Luft et al., 2014).
Таблица 4 . Изменения в сетях мозга при анодной tDCS.
Изменения в сетях мозга при анодной tDCS.
Механизм действия tDCS посредством долговременной потенциации и глиальных клеток
Долговременная потенциация (LTP), непрерывное усиление передачи сигнала между нейронами, как полагают, опосредует эффект tDCS (рис. 3). Во-первых, потенциалы действия в пресинаптических нейронах преобразуются в химические сигналы на пресинаптической мембране. В последующем в синаптическую щель выбрасываются нейротрансмиттеры (глутамат, ГАМК, дофамин, серотонин, ацетилхолин и др.). Процесс, посредством которого этот нейротрансмиттер передается постсинаптическим нейронам, называется каскадом передачи сигнала. В этом каскаде различные нейротрансмиттеры активируют/ингибируют каскады трансдукции, связанные с G-белками или ионными каналами, что приводит к фосфорилированию цАМФ-зависимого белка, связывающего элемент (CREB), и активации генов в ядрах нейронов. Кроме того, каскад трансдукции, связанный с нейротрофическим фактором, может играть роль, активируя различные киназные ферменты (Stephen, 2013). Эти каскады передачи сигнала усиливают синтез различных белков, таких как синтазы нейротрансмиттеров, рецепторы, ионные каналы и внутриклеточные сигнальные белки. Облегчающие действия этих белков, которые регулируют эффективность нейротрансмиссий в цепи коры головного мозга, могут объяснить способность tDCS индуцировать LTP (рис. 3).
Эти каскады передачи сигнала усиливают синтез различных белков, таких как синтазы нейротрансмиттеров, рецепторы, ионные каналы и внутриклеточные сигнальные белки. Облегчающие действия этих белков, которые регулируют эффективность нейротрансмиссий в цепи коры головного мозга, могут объяснить способность tDCS индуцировать LTP (рис. 3).
Рисунок 3 . Предполагаемые механизмы усиления долгосрочного потенцирования tDCS. Различные нейротрансмиттеры активируют/ингибируют каскады трансдукции, связанные с G-белками или ионными каналами, что приводит к фосфорилированию цАМФ-зависимого белка, связывающего элемент (CREB), и активации генов в ядрах нейронов. Эти каскады передачи сигнала усиливают синтез различных белков, таких как синтазы нейротрансмиттеров, рецепторы, ионные каналы и внутриклеточные сигнальные белки. Облегчающие действия этих белков, которые регулируют эффективность нейротрансмиссий в цепи коры головного мозга, могут объяснить способность tDCS индуцировать LTP.
Нейротрофический фактор головного мозга (BDNF) также может опосредовать развитие LTP (Cocco et al., 2018). До сих пор было показано, что многосеансовое анодное моделирование левой дорсолатеральной префронтальной коры (DLPFC) улучшает симптомы настроения без значительного изменения концентрации BDNF в крови пациентов с большим депрессивным расстройством (Brunoni et al., 2015). Необходимы дальнейшие исследования, чтобы увидеть, влияет ли tDCS на уровни BDNF при других психических расстройствах.
Сообщалось, что глиальные клетки, включая астроциты, активируются tDCS (Ruohonen and Karhu, 2012). Поскольку эти клетки регулируют концентрацию химических веществ и нейротрансмиттеров во внешнем пространстве нейронов, механизмы, с помощью которых tDCS улучшает психиатрические симптомы, могут включать некоторые модальности, отличные от прямого воздействия на нейронные клетки. Например, результаты исследований на животных предполагают участие LTP и глиальных клеток (см. Таблицу 5). Сообщалось, что у крыс анодная стимуляция tDCS на синапсах CA3-CA1 гиппокампа индуцирует LTP (Ranieri et al., 2012). Кроме того, tDCS увеличивает накопление цАМФ в поляризованной коре (Hattori et al., 19).90) и изменяет экспрессию мРНК, что приводит к увеличению плотности дендритных шипиков у пациентов с инсультом (Jiang et al., 2012).
Сообщалось, что у крыс анодная стимуляция tDCS на синапсах CA3-CA1 гиппокампа индуцирует LTP (Ranieri et al., 2012). Кроме того, tDCS увеличивает накопление цАМФ в поляризованной коре (Hattori et al., 19).90) и изменяет экспрессию мРНК, что приводит к увеличению плотности дендритных шипиков у пациентов с инсультом (Jiang et al., 2012).
Таблица 5 . Исследования на животных механизма долговременного потенцирования (LTP) анодной tDCS.
Монай и др. сообщили, что tDCS увеличивает уровни норадреналина за счет увеличения внутриклеточных концентраций ионов кальция посредством стимуляции адренергических рецепторов α1 на астроцитах у генетически модифицированных мышей (Monai et al., 2016). Кроме того, в клетках человека было обнаружено увеличение внутриклеточных концентраций ионов кальция с помощью tDCS (Dubé et al., 2012), что свидетельствует об участии астроцитов в способности tDCS индуцировать LTP.
BDNF связывается с рецепторами TrkB, которые регулируют рост и синаптическую активность нейронов и, как полагают, участвуют в формировании LTP (Stephen, 2013). Напр., анодная tDCS индуцирует синаптическую пластичность in vitro , которая зависит от усиленной секреции BDNF и активации TrkB (Fritsch et al., 2010). Более того, Подда и соавт. (2016) сообщили, что у мышей, подвергшихся анодной tDCS, наблюдалась LTP гиппокампа и улучшение обучения и памяти. Сообщалось, что эти эффекты связаны с усилением ацетилирования промотора I BDNF, экспрессией экзонов I и IX BDNF и уровнями белка BDNF (Podda et al., 2016). В гиппокампе мышей, получавших tDCS, также наблюдается повышенное фосфорилирование CREB и фосфорилирование CREB по Ser133 (pCREB 133 ) связывается с промотором I BDNF и рекрутирует CREB-связывающие белки. Эти данные свидетельствуют о том, что анодная tDCS увеличивает LTP гиппокампа и память посредством механизмов, связанных с генами BDNF (Podda et al., 2016; Yu et al., 2019).
Напр., анодная tDCS индуцирует синаптическую пластичность in vitro , которая зависит от усиленной секреции BDNF и активации TrkB (Fritsch et al., 2010). Более того, Подда и соавт. (2016) сообщили, что у мышей, подвергшихся анодной tDCS, наблюдалась LTP гиппокампа и улучшение обучения и памяти. Сообщалось, что эти эффекты связаны с усилением ацетилирования промотора I BDNF, экспрессией экзонов I и IX BDNF и уровнями белка BDNF (Podda et al., 2016). В гиппокампе мышей, получавших tDCS, также наблюдается повышенное фосфорилирование CREB и фосфорилирование CREB по Ser133 (pCREB 133 ) связывается с промотором I BDNF и рекрутирует CREB-связывающие белки. Эти данные свидетельствуют о том, что анодная tDCS увеличивает LTP гиппокампа и память посредством механизмов, связанных с генами BDNF (Podda et al., 2016; Yu et al., 2019).
Выводы
В этом обзоре мы обсудили электрофизиологическое понимание tDCS на основе модели, зависящей от стимуляции. Биохимически усиление возбуждающих синаптических передач посредством анодной стимуляции, вероятно, способствует передаче глутамата и подавляет передачу гамма-аминомасляной кислоты в коре.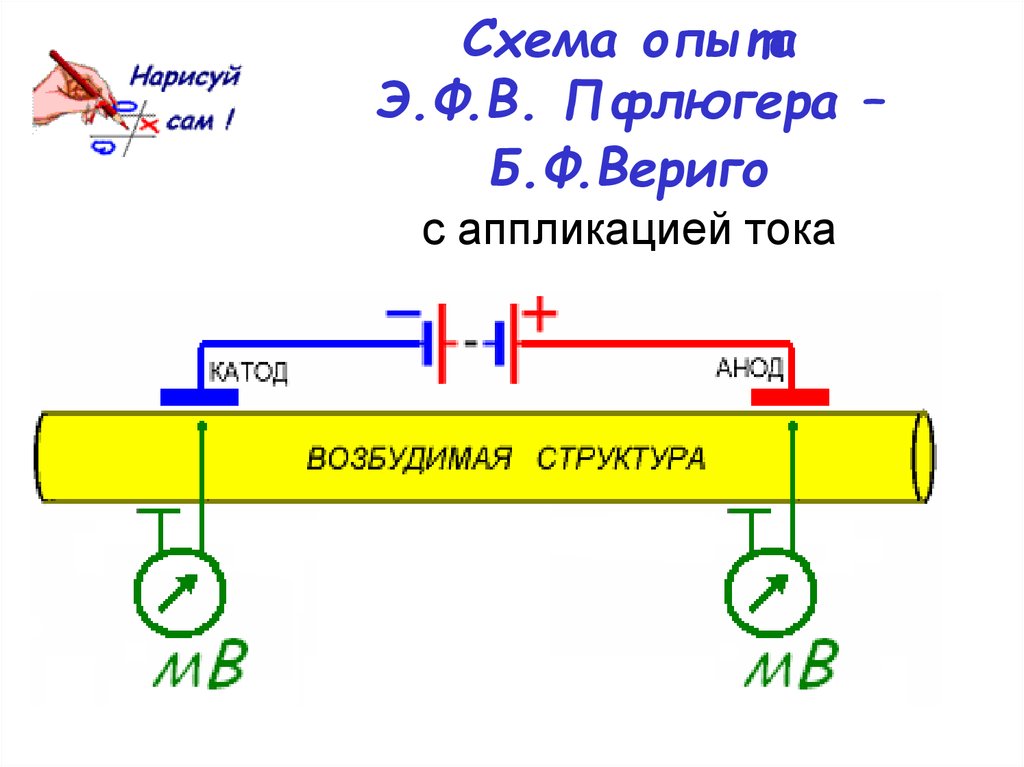 Соответственно, tDCS может положительно или отрицательно регулировать передачу дофамина, серотонина и ацетилхолина. Эти нейронные события могут изменить баланс между возбуждающими и тормозными входами. Таким образом, анодная стимуляция может модулировать уровни активности нескольких сетевых систем.
Соответственно, tDCS может положительно или отрицательно регулировать передачу дофамина, серотонина и ацетилхолина. Эти нейронные события могут изменить баланс между возбуждающими и тормозными входами. Таким образом, анодная стимуляция может модулировать уровни активности нескольких сетевых систем.
LTP может также обеспечивать предполагаемые механизмы, лежащие в основе способности tDCS лечить психические расстройства. В будущих исследованиях следует рассмотреть другие области симптомов психических состояний шизофрении и расстройств настроения, например, социальное познание и метапознание (Nishida et al., 2018; Yamada et al., 2019), в качестве цели лечения tDCS. Кроме того, идентификация предикторов его терапевтических преимуществ в клинических условиях заслуживает дальнейших усилий (Bulubas et al., 2019).
Ограничения
Текущий обзор является описательным, и статьи не подвергались систематическому поиску. Более того, многие статьи, представленные в этом обзоре, были нацелены на здоровых людей, а не на людей с психическими расстройствами. Следует отметить, что пациенты с психическими заболеваниями могут иначе реагировать на tDCS, чем здоровые люди. Кроме того, некоторые статьи, включенные в этот обзор, были посвящены влиянию tDCS на моторную кору. Необходимы дальнейшие исследования для изучения доказательств стимуляции префронтальной коры, которая была основной мишенью психических расстройств, таких как депрессия и шизофрения (Mezger et al., 2020).
Следует отметить, что пациенты с психическими заболеваниями могут иначе реагировать на tDCS, чем здоровые люди. Кроме того, некоторые статьи, включенные в этот обзор, были посвящены влиянию tDCS на моторную кору. Необходимы дальнейшие исследования для изучения доказательств стимуляции префронтальной коры, которая была основной мишенью психических расстройств, таких как депрессия и шизофрения (Mezger et al., 2020).
Вклад авторов
YY и TS спланировали и разработали обзор, внесли существенный вклад и одобрили окончательный вариант рукописи. YY собрал данные и составил первую рукопись. Т.С. критически рассмотрел проект и отредактировал его. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Это исследование было поддержано Японским обществом содействия развитию науки (JSPS) KAKENHI № 20K16635, внутренний исследовательский грант (29-1, 30-1 и 30-8) для неврологических и психических расстройств Национального центра неврологии и психиатрии (NCNP) и JH 2020-B-08, а также AMED под номерами грантов 18dk0307069и 18дк0307081.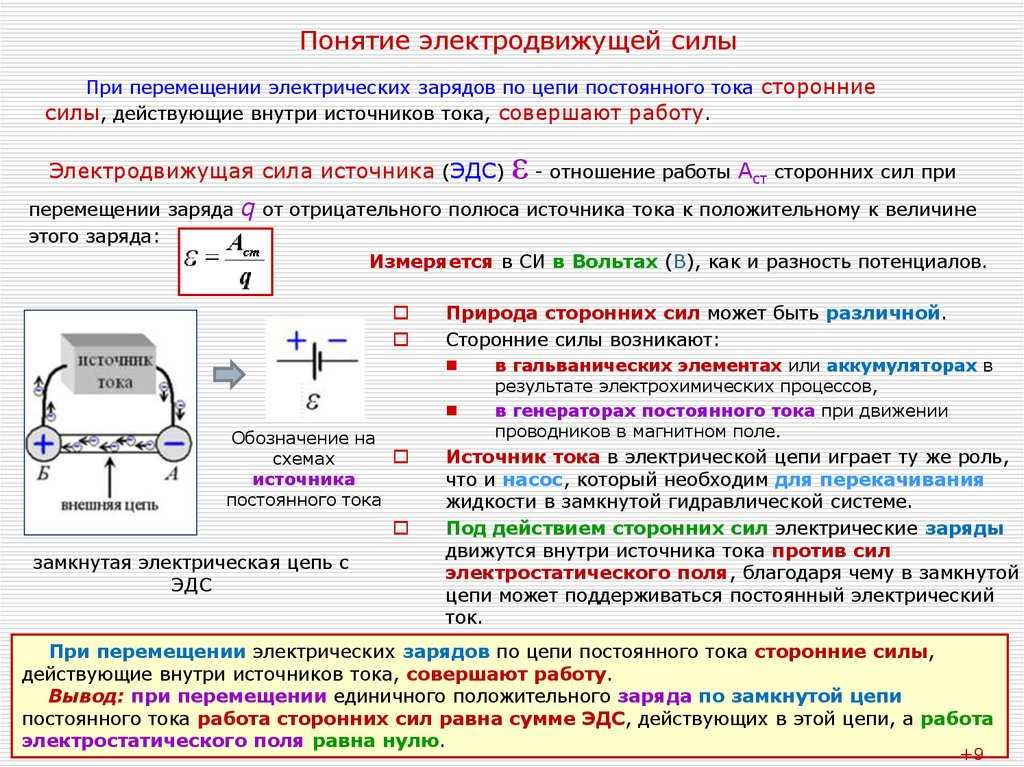
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы хотели бы поблагодарить Drs. Казуюки Накагоме, Шинсуке Кито и Наоцугу Хирабаяши из Национального центра неврологии и психиатрии за поддержку нашей исследовательской деятельности.
Ссылки
Bachtiar, V., Near, J., Johansen-Berg, H., and Stagg, C.J. (2015). Модуляция функциональной связи ГАМК и состояния покоя путем транскраниальной стимуляции постоянным током. Элиф 4:e08789. doi: 10.7554/eLife.08789
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Брунони А. Р., Баекен К., Мачадо-Виейра Р., Гаттаз В. Ф. и Вандерхасселт М. А. (2015). Уровни BDNF в крови после неинвазивных вмешательств по стимуляции мозга при большом депрессивном расстройстве: систематический обзор и метаанализ.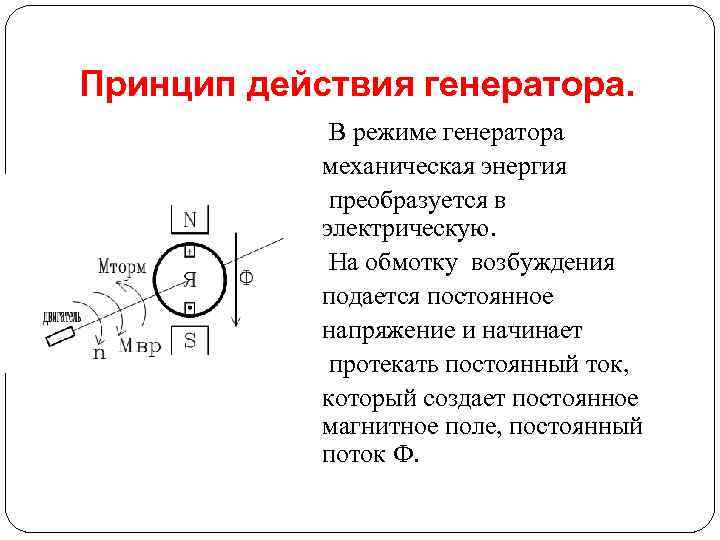 World J. Biol. Психиатрия 16, 114–122. doi: 10.3109/15622975.2014.958101
World J. Biol. Психиатрия 16, 114–122. doi: 10.3109/15622975.2014.958101
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Булубас Л., Падберг Ф., Буэно П.В., Дюран Ф., Бусатто Г., Амаро Э. мл. и др. (2019). Антидепрессивные эффекты tDCS связаны с префронтальными объемами серого вещества в начале исследования: данные исследования ELECT-TDCS. Стимуляция мозга . 12, 1197–1204. doi: 10.1016/j.brs.2019.05.006
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Chai, X. J., Whitfield-Gabrieli, S., Shinn, A. K., Gabrieli, J. D., Nieto Castañón, A., McCarthy, J. M., et al. (2011). Аномальная связность медиальной префронтальной коры в состоянии покоя при биполярном расстройстве и шизофрении. Нейропсихофармакология 36, 2009–2017. doi: 10.1038/npp.2011.88
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Чайеб Л., Антал А., Терней Д. и Паулюс В. (2012). Фармакологическая модуляция кратковременных эффектов антагонистической стимуляции постоянным током моторной коры человека. Фронт. Психиатрия 3:67. doi: 10.3389/fpsyt.2012.00067
Фронт. Психиатрия 3:67. doi: 10.3389/fpsyt.2012.00067
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Кларк В. П., Коффман Б. А., Трамбо М. К. и Гаспарович К. (2011). Транскраниальная стимуляция постоянным током (tDCS) вызывает локальные и специфические изменения в нейрохимии:0017 1 H Исследование методом магнитно-резонансной спектроскопии. Неврологи. Письмо . 500, 67–71. doi: 10.1016/j.neulet.2011.05.244
CrossRef Полный текст | Google Scholar
Кокко С., Подда М. В. и Грасси К. (2018). Роль передачи сигналов BDNF в улучшении памяти, вызванном транскраниальной стимуляцией постоянным током. Фронт. Нейроски . 12:427. doi: 10.3389/fnins.2018.00427
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Du, Y., Pearlson, G.D., Lin, D., Sui, J., Chen, J., Salman, M., et al. (2017). Идентификация биомаркеров динамической функциональной связи с использованием GIG-ICA: применение к шизофрении, шизоаффективному расстройству и психотическому биполярному расстройству. Гул. Карта мозга . 38, 2683–2708. doi: 10.1002/hbm.23553
Гул. Карта мозга . 38, 2683–2708. doi: 10.1002/hbm.23553
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Дюбе Дж., Рошетт-Друэн О., Левеск П., Говен Р., Роберж С. Дж., Оже Ф. А. и др. (2012). Кератиноциты человека реагируют на стимуляцию постоянным током, увеличивая внутриклеточный кальций: преимущественная реакция низкодифференцированных клеток. Дж. Сотовый. Физиол. 227, 2660–2667. doi: 10.1002/jcp.23008
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Фертонани, А., Пирулли, К., и Миниусси, К. (2011). Стимуляция случайным шумом улучшает нейропластичность в перцептивном обучении. Дж. Нейроски . 31, 15416–15423. doi: 10.1523/JNEUROSCI.2002-11.2011
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Фонтено К., Редуте Дж., Хазебарт Ф., Ле Барс Д., Кост Н., Суо-Шаньи М. Ф. и др. (2018). Фронтальная транскраниальная стимуляция постоянным током вызывает высвобождение дофамина в вентральном стриатуме у человека. Церебр. Кортекс . 28, 2636–2646. doi: 10.1093/cercor/bhy093
Церебр. Кортекс . 28, 2636–2646. doi: 10.1093/cercor/bhy093
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Фрегни Ф., Эль-Хаграсси М. М., Пачеко-Барриос К., Карвалью С., Лейте Дж., Симис М. и др. (2020). Основанные на фактических данных рекомендации и вторичный метаанализ по использованию транскраниальной стимуляции постоянным током (tDCS) при неврологических и психических расстройствах. Междунар. J. Нейропсихофармакол. пяа051. дои: 10.1093/ijnp/pyaa051. [Epub перед печатью].
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Фрич Б., Рейс Дж., Мартинович К., Шамбра Х. М., Джи Ю., Коэн Л. Г. и др. (2010). Стимуляция постоянным током способствует BDNF-зависимой синаптической пластичности: потенциальные последствия для моторного обучения. Нейрон 66, 198–204. doi: 10.1016/j.neuron.2010.03.035
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Фукаи М., Бунаи Т.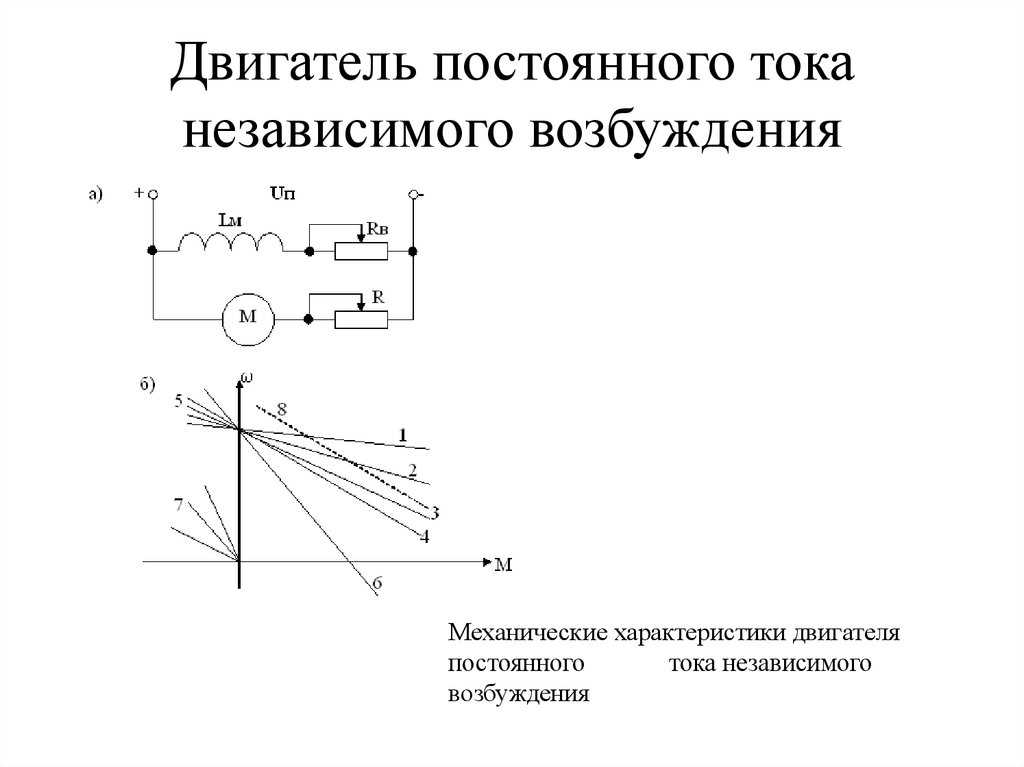 , Хиросава Т., Кикучи М., Ито С., Минабэ Ю. и др. (2019). Высвобождение эндогенного дофамина при транскраниальной стимуляции постоянным током регулирует повышенное внимание: исследование с позитронно-эмиссионной томографией. Перевод. Психиатрия 9:115. doi: 10.1038/s41398-019-0443-4
, Хиросава Т., Кикучи М., Ито С., Минабэ Ю. и др. (2019). Высвобождение эндогенного дофамина при транскраниальной стимуляции постоянным током регулирует повышенное внимание: исследование с позитронно-эмиссионной томографией. Перевод. Психиатрия 9:115. doi: 10.1038/s41398-019-0443-4
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Хаттори Ю., Мориваки А. и Хори Ю. (1990). Двухфазные эффекты поляризующего тока на чувствительную к аденозину генерацию циклического АМФ в коре головного мозга крыс. Неврологи. Письмо . 116, 320–324. дои: 10.1016/0304-3940(90)-P
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Хантер, М. А., Коффман, Б. А., Гаспарович, К., Калхун, В. Д., Трамбо, М. К., и Кларк, В. П. (2015). Базовые эффекты транскраниальной стимуляции постоянным током на глутаматергическую нейротрансмиссию и крупномасштабную сетевую связь. Мозг Res . 1594, 92–107. doi: 10.1016/j.brainres.2014.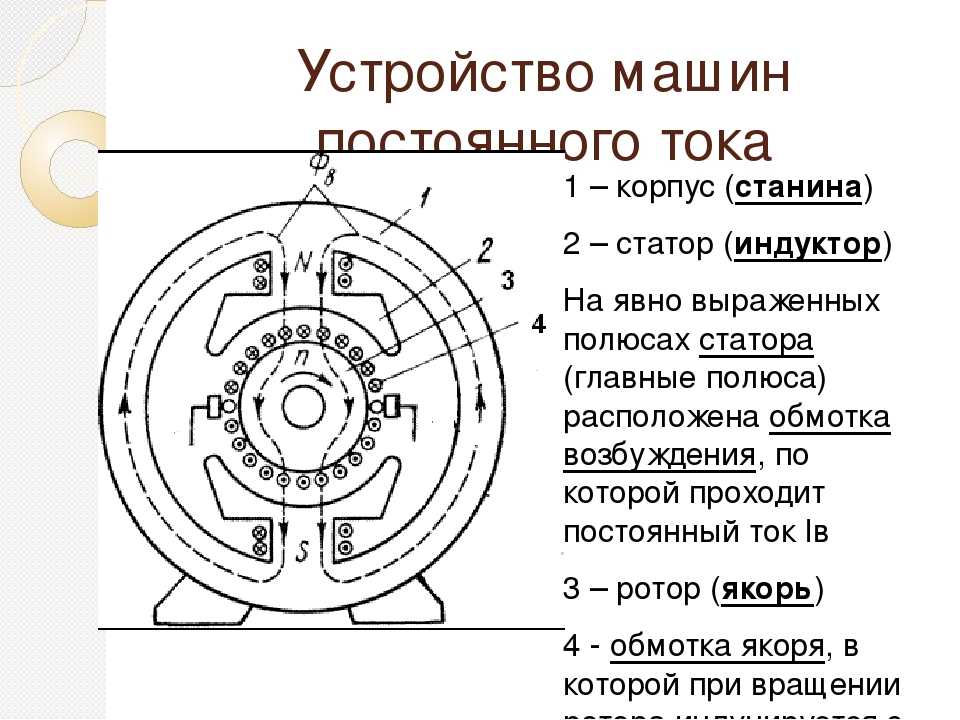 09.066
09.066
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Jiang, T., Xu, R. X., Zhang, A. W., Di, W., Xiao, Z. J., Miao, J. Y., et al. (2012). Влияние транскраниальной стимуляции постоянным током на паннексин-1 гемиканала и нейронную пластичность в крысиной модели инфаркта головного мозга. Неврология 226, 421–426. doi: 10.1016/j.neuroscience.2012.09.035
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ким Дж., Ивата Ю., Плитман Э., Караваджо Ф., Чанг Дж. К., Шах П. и др. (2019). Метаанализ транскраниальной стимуляции постоянным током при шизофрении: «Чем больше, тем лучше?» J. Psychiatr. Рез. 110, 117–126. doi: 10.1016/j.jpsychires.2018.12.009
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Kunze, T., Hunold, A., Haueisen, J., Jirsa, V., and Spiegler, A. (2016). Транскраниальная стимуляция постоянным током изменяет функциональную связность в состоянии покоя: крупномасштабное исследование моделирования сети мозга.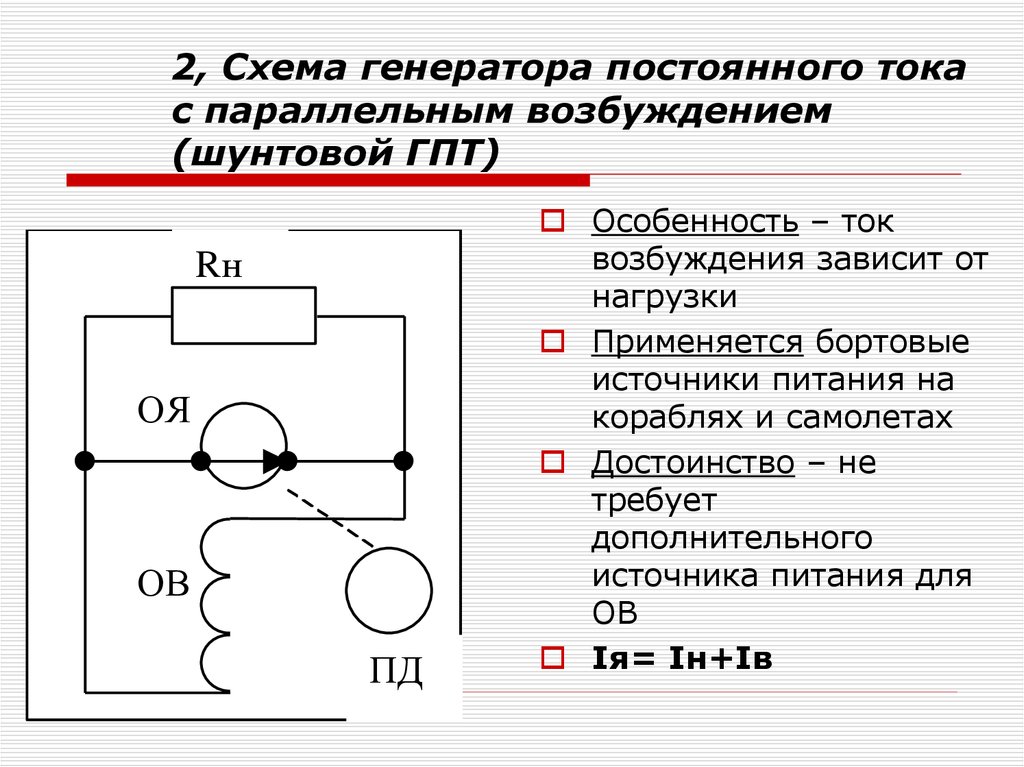 Нейроизображение 140, 174–187. doi: 10.1016/j.neuroimage.2016.02.015
Нейроизображение 140, 174–187. doi: 10.1016/j.neuroimage.2016.02.015
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Куо М.Ф., Грош Дж., Фрегни Ф., Паулюс В. и Ницше М.А. (2007). Фокусирующее влияние ацетилхолина на нейропластичность моторной коры человека. Дж. Нейроски . 27, 14442–14447. doi: 10.1523/JNEUROSCI.4104-07.2007
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Куо, М. Ф., Паулюс, В., и Ницше, М. А. (2008). Повышение очагово-индуцированной пластичности мозга дофамином. Церебр. Кортекс . 18, 648–651. doi: 10.1093/cercor/bhm098
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Либетанц Д., Ницше М.А., Тергау Ф. и Паулюс В. (2002). Фармакологический подход к механизмам последействия транскраниальной стимуляции постоянным током возбудимости моторной коры человека. Мозг . 125 (часть 10), 2238–2247. doi: 10.1093/brain/awf238
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Люфт, К. Д., Переда, Э., Банисси, М. Дж., и Бхаттачарья, Дж. (2014). Лучшее из обоих миров: обещание сочетать стимуляцию мозга и коннектом мозга. Фронт. Сист. Нейроски . 8:132. doi: 10.3389/fnsys.2014.00132
Д., Переда, Э., Банисси, М. Дж., и Бхаттачарья, Дж. (2014). Лучшее из обоих миров: обещание сочетать стимуляцию мозга и коннектом мозга. Фронт. Сист. Нейроски . 8:132. doi: 10.3389/fnsys.2014.00132
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Медейрос Л. Ф., де Соуза И. К., Видор Л. П., де Соуза А., Дейтос А., Волц М. С. и др. (2012). Нейробиологические эффекты транскраниальной стимуляции постоянным током: обзор. Фронт. Психиатрия 3:110. doi: 10.3389/fpsyt.2012.00110
PubMed Abstract | CrossRef Full Text
Мерзагора А.С., Фоффани Г., Паньявин И., Мордильо-Матеос Л., Агилар Дж., Онарал Б. и др. (2010). Префронтальные гемодинамические изменения, вызванные анодной стимуляцией постоянным током. Нейроизображение 49, 2304–2310. doi: 10.1016/j.neuroimage.2009.10.044
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Мезгер Э., Раухманн Б. С., Брунони А. Р., Булубас Л., Тильшер А.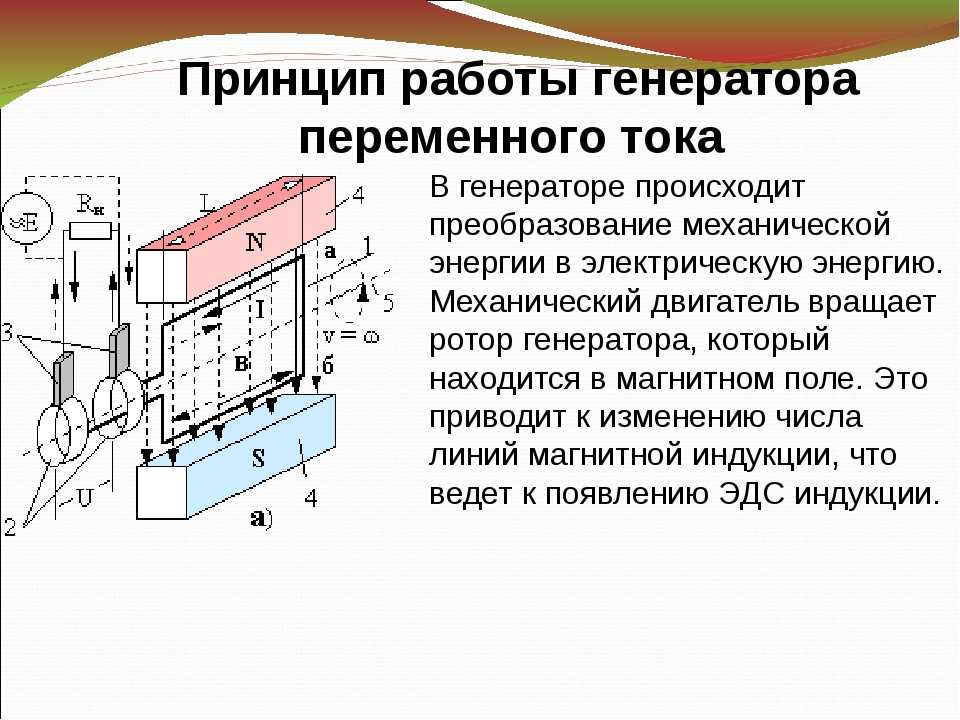 , Верле Дж. и др. (2020). Влияние бифронтальной транскраниальной стимуляции постоянным током на уровни глутамата в головном мозге и связь в состоянии покоя: мультимодальные данные МРТ для участка катодной стимуляции. евро. Арка Психиатрия клин. Нейроски . doi: 10.1007/s00406-020-01177-0. [Epub перед печатью].
, Верле Дж. и др. (2020). Влияние бифронтальной транскраниальной стимуляции постоянным током на уровни глутамата в головном мозге и связь в состоянии покоя: мультимодальные данные МРТ для участка катодной стимуляции. евро. Арка Психиатрия клин. Нейроски . doi: 10.1007/s00406-020-01177-0. [Epub перед печатью].
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Monai, H., Okura, M., Tanaka, M., Oe, Y., Konno, A., Hirai, H., et al. (2016). Визуализация кальция показывает участие глии в пластичности, индуцированной транскраниальной стимуляцией постоянным током, в мозге мышей. Нац. Коммуна . 7:11100. doi: 10.1038/ncomms11100
PubMed Abstract | Полный текст CrossRef
Монте-Сильва, К., Либетанц, Д., Грандей, Дж., Паулюс, В., и Ницше, М.А. (2010). Нелинейный эффект леводопы в зависимости от дозировки на пластичность моторной коры человека. J. Physiol. 588 (часть 18), 3415–3424. doi: 10.1113/jphysiol.2010.1
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Нарита З. , Стикли А., ДеВилдер Дж., Ёкои Ю., Инагава Т., Ямада Ю. и др. (2020). Влияние мультисессионной префронтальной транскраниальной стимуляции постоянным током на когнитивные функции при шизофрении: систематический обзор и метаанализ. Шизофр. Рез . 216, 367–373. doi: 10.1016/j.schres.2019.11.011
, Стикли А., ДеВилдер Дж., Ёкои Ю., Инагава Т., Ямада Ю. и др. (2020). Влияние мультисессионной префронтальной транскраниальной стимуляции постоянным током на когнитивные функции при шизофрении: систематический обзор и метаанализ. Шизофр. Рез . 216, 367–373. doi: 10.1016/j.schres.2019.11.011
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Нисида К., Тойомаки А., Кошикава Ю., Ниимура Х., Моримото Т., Тани М. и др. (2018). Социальное познание и метапознание способствуют точности самооценки функционирования в реальном мире у пациентов с шизофренией. Шизофр. Рез . 202, 426–428. doi: 10.1016/j.schres.2018.06.071
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Нитше М.А., Фрике К., Хеншке У., Шлиттерлау А., Либетанц Д., Ланг Н. и др. (2003). Фармакологическая модуляция сдвигов возбудимости коры, вызванных транскраниальной стимуляцией постоянным током у человека. J Физиол. 553 (часть 1), 293–301. doi: 10.1113/jphysiol.2003.049916
doi: 10.1113/jphysiol.2003.049916
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ницше, М. А., Гранди, Дж., Либетанц, Д., Ланг, Н., Тергау, Ф., и Паулюс, В. (2004a). Катехоламинергическая консолидация нейропластичности моторной коры у человека. Церебр. Кортекс . 14, 1240–1245. doi: 10.1093/cercor/bhh085
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ницше М.А., Яусси В., Либетанц Д., Ланг Н., Тергау Ф. и Паулюс В. (2004b). Консолидация моторной корковой нейропластичности человека с помощью D-циклосерина. Нейропсихофармакология 29, 1573–1578. doi: 10.1038/sj.npp.1300517
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ницше М.А., Куо М.Ф., Карраш Р., Вехтер Б., Либетанц Д. и Паулюс В. (2009 г.)). Серотонин влияет на транскраниальную нейропластичность, индуцированную постоянным током, у людей. Биол. Психиатрия 66, 503–508. doi: 10.1016/j.biopsych.2009. 03.022
03.022
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Нитше М. А., Лампе К., Антал А., Либетанц Д., Ланг Н., Тергау Ф. и др. (2006). Дофаминергическая модуляция длительных изменений возбудимости коры, индуцированных постоянным током, в моторной коре человека. евро. Дж. Нейроски . 23, 1651–1657. дои: 10.1111/j.1460-9568.2006.04676.x
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Нитше, М. А., Либетанц, Д., Шлиттерлау, А., Хеншке, У., Фрике, К., Фромманн, К., и др. (2004с). ГАМКергическая модуляция сдвигов возбудимости моторной коры, вызванных стимуляцией постоянного тока, у людей. евро. Дж. Нейроски . 19, 2720–2726. doi: 10.1111/j.0953-816X.2004.03398.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ницше, М. А., и Паулюс, В. (2000). Изменения возбудимости, вызванные в моторной коре человека слабой транскраниальной стимуляцией постоянным током. J Физиол. 527 (часть 3), 633–639. doi: 10.1111/j.1469-7793.2000.t01-1-00633.x
527 (часть 3), 633–639. doi: 10.1111/j.1469-7793.2000.t01-1-00633.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Окун, М., и Лэмпл, И. (2008). Мгновенная корреляция возбуждения и торможения при текущей и сенсорно-вызванной деятельности. Нац. Нейроски . 11, 535–537. doi: 10.1038/nn.2105
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Филип Н. С., Нельсон Б. Г., Фролих Ф., Лим К. О., Видж А. С. и Карпентер Л. Л. (2017). Стимуляция низкоинтенсивным транскраниальным током в психиатрии. утра. Журнал психиатрии 174, 628–639. doi: 10.1176/appi.ajp.2017.160
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Подда М.В., Кокко С., Мастродонато А., Фуско С., Леоне Л., Барбати С.А. и др. (2016). Анодальная транскраниальная стимуляция постоянным током повышает синаптическую пластичность и память у мышей посредством эпигенетической регуляции экспрессии Bdnf. наук. Реп . 6:22180. doi: 10.1038/srep22180
Реп . 6:22180. doi: 10.1038/srep22180
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Полания, Р., Паулюс, В., Антал, А., и Ницше, М.А. (2011). Введение теории графов для отслеживания нейропластических изменений в мозге человека в состоянии покоя: исследование транскраниальной стимуляции постоянным током. Нейроизображение 54, 2287–2296. doi: 10.1016/j.neuroimage.2010.09.085
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ранго М., Коджаманян Ф., Марселья С., Барберис Б., Ариги А., Биондетти П. и др. (2008). Содержание миоинозитола в человеческом мозгу изменяется транскраниальной стимуляцией постоянным током за считанные минуты: исследование 1H-MRS. Маг. Резон. Мед . 60, 782–789. doi: 10.1002/mrm.21709
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ranieri, F., Podda, M.V., Riccardi, E., Frisullo, G., Dileone, M., Profice, P., et al. (2012). Модуляция LTP в синапсах CA3-CA1 гиппокампа крысы путем стимуляции постоянным током. Дж. Нейрофизиол . 107, 1868–1880 гг. doi: 10.1152/jn.00319.2011
Дж. Нейрофизиол . 107, 1868–1880 гг. doi: 10.1152/jn.00319.2011
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Руохонен, Дж., и Карху, Дж. (2012). tDCS, возможно, стимулирует глиальные клетки. клин. Нейрофизиол . 123, 2006–2009 гг. doi: 10.1016/j.clinph.2012.02.082
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Сео Х. и Джун С.К. (2019). Связь между электрическим полем и активацией корковых нейронов при транскраниальной электрической стимуляции. Стимуляция мозга . 12, 275–289. doi: 10.1016/j.brs.2018.11.004
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Шиодзава П., Фрегни Ф., Бенсеньор И. М., Лотуфо П. А., Берлим М. Т. и Даскалакис Дж. З. (2014). Транскраниальная стимуляция постоянным током при большой депрессии: обновленный систематический обзор и метаанализ. Междунар. Дж. Нейропсихофармакол . 17, 1443–1452. doi: 10.1017/S1461145714000418
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Сильванто Дж.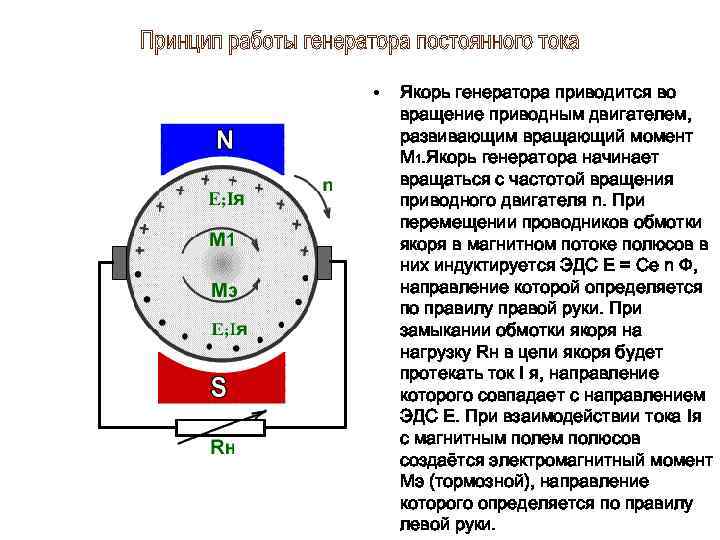 , Магглтон Н. и Уолш В. (2008). Зависимость от состояния в исследованиях восприятия и познания со стимуляцией мозга. Тенденции Cogn. науч. 12, 447–454. doi: 10.1016/j.tics.2008.09.004
, Магглтон Н. и Уолш В. (2008). Зависимость от состояния в исследованиях восприятия и познания со стимуляцией мозга. Тенденции Cogn. науч. 12, 447–454. doi: 10.1016/j.tics.2008.09.004
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Stagg, C.J., Bachtiar, V., Amadi, U., Gudberg, C.A., Ilie, A.S., Sampaio-Baptista, C., et al. (2014). Локальная концентрация ГАМК связана с функциональной связностью в состоянии покоя на уровне сети. Элиф 3:e01465. doi: 10.7554/eLife.01465
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Stagg, CJ, Bachtiar, V., and Johansen-Berg, H. (2011). Роль ГАМК в моторном обучении человека. Курс. Биол . 21, 480–484. doi: 10.1016/j.cub.2011.01.069
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Stagg, C.J., Best, JG, Stephenson, M.C., O’Shea, J., Wylezinska, M., Kincses, Z.T., et al. (2009). Чувствительная к полярности модуляция корковых нейротрансмиттеров при транскраниальной стимуляции. Дж. Нейроски . 29, 5202–5206. doi: 10.1523/JNEUROSCI.4432-08.2009
Дж. Нейроски . 29, 5202–5206. doi: 10.1523/JNEUROSCI.4432-08.2009
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Stagg, CJ, and Nitsche, MA (2011). Физиологические основы транскраниальной стимуляции постоянным током. Невролог 17, 37–53. doi: 10.1177/1073858410386614
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Стивен М.С. (2013). Основная психофармакология Шталя: нейронаучные основы и практические приложения, 4-е изд., . Нью-Йорк, штат Нью-Йорк: Издательство Кембриджского университета.
Google Scholar
Тиругнансамбандам Н., Гранди Дж., Адам К., Дрис А., Сквирба А.С., Ланг Н. и др. (2011). Никотинергическое влияние на очаговую и неочаговую нейропластичность, вызванную неинвазивной стимуляцией мозга у некурящих людей. Нейропсихофармакология 36, 879–886. doi: 10.1038/npp.2010.227
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Вершинг Дж.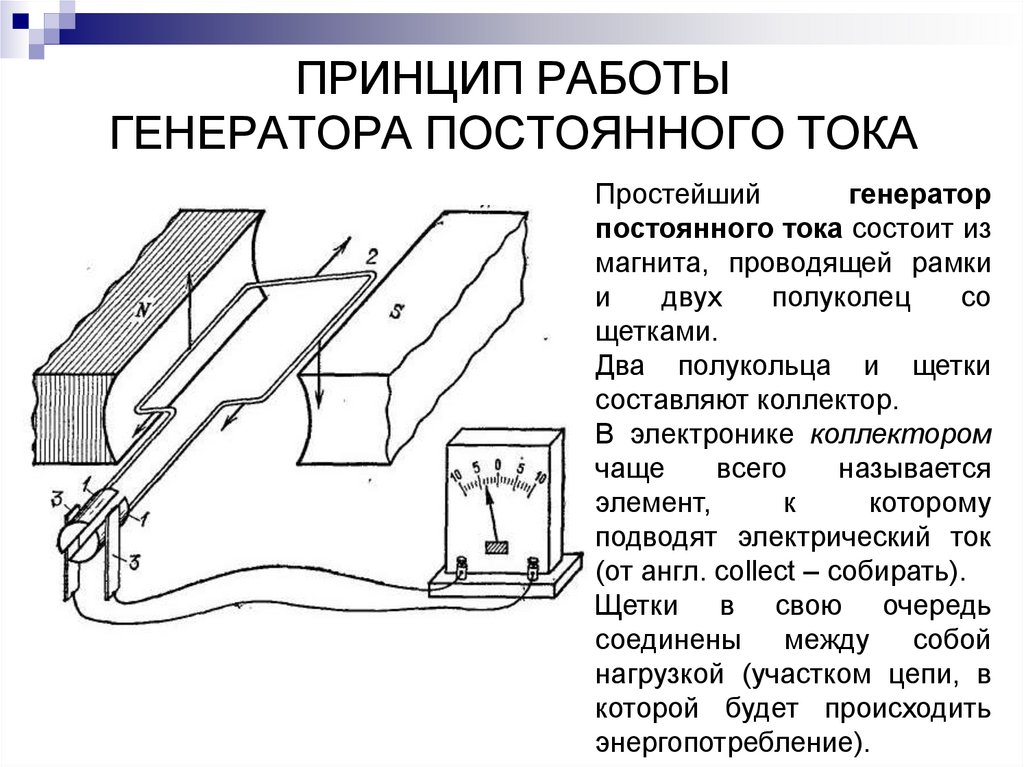 , Падберг Ф., Геригк С., Хайнц И., Бауэр К., Плевниа К. и др. (2018). Проверка предположений о префронтальной транскраниальной стимуляции постоянным током: сравнение электродных монтажей с использованием мультимодальной фМРТ. Стимуляция мозга . 11, 998–1007. doi: 10.1016/j.brs.2018.05.001
, Падберг Ф., Геригк С., Хайнц И., Бауэр К., Плевниа К. и др. (2018). Проверка предположений о префронтальной транскраниальной стимуляции постоянным током: сравнение электродных монтажей с использованием мультимодальной фМРТ. Стимуляция мозга . 11, 998–1007. doi: 10.1016/j.brs.2018.05.001
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Yamada, Y., Inagawa, T., Sueyoshi, K., Sugawara, N., Ueda, N., Omachi, Y., et al. (2019). Дефицит социального познания как цель раннего вмешательства при психозах: систематический обзор. Фронт. Психиатрия 10:333. doi: 10.3389/fpsyt.2019.00333
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ямада Ю., Мацумото М., Иидзима К. и Сумиёси Т. (2020). Специфичность и преемственность шизофрении и биполярного расстройства: связь с биомаркерами. Курс. фарм. Дез . 26, 191–200. doi: 10.2174/13816128256661
153508
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ёкои Ю. , Нарита З. и Сумиёси Т. (2018). Транскраниальная стимуляция постоянным током при депрессии и психозе: систематический обзор. клин. ЭЭГ Neurosci . 49, 93–102. doi: 10.1177/1550059417732247
, Нарита З. и Сумиёси Т. (2018). Транскраниальная стимуляция постоянным током при депрессии и психозе: систематический обзор. клин. ЭЭГ Neurosci . 49, 93–102. doi: 10.1177/1550059417732247
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Ю, Т. Х., Ву, Ю. Дж., Чиен, М. Э., и Хсу, К. С. (2019 г.)). Транскраниальная стимуляция постоянным током вызывает метапластичность гиппокампа, опосредованную мозговым нейротрофическим фактором. Нейрофармакология 144, 358–367. doi: 10.1016/j.neuropharm.2018.11.012
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Что такое транскраниальная стимуляция постоянным током (tDCS)?
Основы транскраниальной стимуляции постоянным током (tDCS):
Транскраниальная стимуляция постоянным током (tDCS) — это портативный, носимый метод стимуляции мозга, который подает слабый электрический ток на кожу головы. Обычно применяется фиксированный ток от 1 до 2 мА 1 . tDCS работает, подавая положительный (анодный) или отрицательный (катодный) ток через электроды на область. tDCS — это метод нейромодуляции, который вызывает немедленные и длительные изменения в функции мозга. Положение анодного и катодного электродов на голове используется для определения того, как ток течет к определенным областям мозга. Ток, подаваемый tDCS, НЕ достаточно силен, чтобы вызвать потенциал действия в нейроне; вместо этого его «подпорог» изменяет структуру уже активных нейронов. Думайте о мозге как об активном человеке, пытающемся что-то сделать или чему-то научиться, и tDCS, подстегивающем эту постоянную активность. На клеточном уровне 2 , tDCS изменяет возбуждение нейронов и, усиливая синаптическую передачу между нейронами, увеличивает синаптическую пластичность 3 , которая, в свою очередь, является клеточной основой обучения. tDCS часто сочетают с обучением. Тренировка сама по себе производит обучение (синаптическая пластичность), а одновременная tDCS усиливает эти эффекты (повышает синаптическую пластичность).
tDCS работает, подавая положительный (анодный) или отрицательный (катодный) ток через электроды на область. tDCS — это метод нейромодуляции, который вызывает немедленные и длительные изменения в функции мозга. Положение анодного и катодного электродов на голове используется для определения того, как ток течет к определенным областям мозга. Ток, подаваемый tDCS, НЕ достаточно силен, чтобы вызвать потенциал действия в нейроне; вместо этого его «подпорог» изменяет структуру уже активных нейронов. Думайте о мозге как об активном человеке, пытающемся что-то сделать или чему-то научиться, и tDCS, подстегивающем эту постоянную активность. На клеточном уровне 2 , tDCS изменяет возбуждение нейронов и, усиливая синаптическую передачу между нейронами, увеличивает синаптическую пластичность 3 , которая, в свою очередь, является клеточной основой обучения. tDCS часто сочетают с обучением. Тренировка сама по себе производит обучение (синаптическая пластичность), а одновременная tDCS усиливает эти эффекты (повышает синаптическую пластичность). В настоящее время изучаются некоторые клинические применения tDCS: депрессия, шизофрения, афазия, зависимость 4 , эпилепсия, хроническая боль (мигрень, фибромиалгия), внимание и двигательная реабилитация. tDCS также используется для немедицинских оздоровительных приложений, например, для ускоренного обучения 5 , концентрация, расслабление и медитация. 6
В настоящее время изучаются некоторые клинические применения tDCS: депрессия, шизофрения, афазия, зависимость 4 , эпилепсия, хроническая боль (мигрень, фибромиалгия), внимание и двигательная реабилитация. tDCS также используется для немедицинских оздоровительных приложений, например, для ускоренного обучения 5 , концентрация, расслабление и медитация. 6
Как выглядит устройство для транскраниальной стимуляции постоянным током (tDCS)?
Устройства tDCS представляют собой небольшие устройства с батарейным питанием. Обычно имеется панель управления, которая позволяет программировать устройство (устанавливать продолжительность и интенсивность стимуляции). Электроды надеваются на голову и удерживаются головным убором — обычно эластичным ремешком. Кабель соединяет каждый электрод со стимулятором. При включении стимулятора ток идет от устройства к электроду, а затем через мозг. Стимуляторы профессионального уровня имеют множество функций, которые помогают обеспечить переносимость и надежность стимуляции. Это включает в себя измеритель импеданса и измеритель тока.
Это включает в себя измеритель импеданса и измеритель тока.
Каковы побочные эффекты транскраниальной стимуляции постоянным током (tDCS)?
Исследования побочных эффектов tDCS продолжаются, но до сих пор установленные побочные эффекты незначительны 7 и ограничиваются расположением электрода. Они включают временное покраснение кожи, зуд и покалывание. Другие предполагаемые побочные эффекты tDCS включают головную боль, тошноту и головокружение. Следует отметить, что эти последние три побочных эффекта, как было показано, проявляются почти с той же частотой, что и ложная стимуляция (фальшивая стимуляция) 8 При неправильном применении tDCS могут возникнуть другие побочные эффекты, такие как фосфен, представляющий собой временную неопасную вспышку света. Это может произойти, если электроды расположены слишком близко к глазу. Кроме того, неправильное введение tDCS может вызвать стандартные ожоги кожи. Нет никаких научных доказательств, демонстрирующих стойкие травмы или необратимые побочные эффекты от tDCS. Тем не менее, следует отметить, что все данные о переносимости и безопасности tDCS получены в результате контролируемых испытаний на людях с использованием специализированного оборудования и строго контролируемых протоколов (например, ограничение текущей продолжительности, количество сеансов).
Тем не менее, следует отметить, что все данные о переносимости и безопасности tDCS получены в результате контролируемых испытаний на людях с использованием специализированного оборудования и строго контролируемых протоколов (например, ограничение текущей продолжительности, количество сеансов).
На что похожа транскраниальная стимуляция постоянным током (tDCS)?
Во время tDCS большинство людей ощущают легкое покалывание, покалывание, зуд или тепло. Эти ощущения безболезненны и проходят при прекращении стимуляции. Однако «переносимость» tDCS зависит от качества принадлежностей, процедур настройки и использования интенсивности (несколько мА) и продолжительности (десятки минут), соответствующих стандартам tDCS. 23
Чего мы не знаем о транскраниальной стимуляции постоянным током (tDCS)?
В то время как остаются вопросы о лучших приложениях для tDCS, существуют десятилетия исследований, обозначающих его причастный механизм. Недавняя работа предполагает, что активация глии и изменение внутриклеточных концентраций цАМФ и кальция в значительной степени способствуют эффектам tDCS. временная потенциация (ДП) и длительная депрессия (ДД).
временная потенциация (ДП) и длительная депрессия (ДД).
Где можно пройти транскраниальную стимуляцию постоянным током (tDCS)?
В США tDCS имеет регулирующий статус «исследовательского» 10 . Это не указывает на эффективность; это означает, что FDA не выдало заключение. Как правило, FDA не дает заключения до тех пор, пока компании не проявят интерес к маркетингу устройства. В Соединенных Штатах компаниям не разрешается продавать tDCS по клиническим показаниям, таким как «лечение депрессии» или «лечение эпилепсии». Врачам в Соединенных Штатах разрешено проводить лечение «не по прямому назначению», то есть лечение, которое не одобрено FDA для данного показания. Исследовательским центрам по всему миру разрешено тестировать tDCS в контролируемых клинических испытаниях. В таких испытаниях каждый субъект должен подписать лист информированного согласия. Вы можете найти список испытаний tDCS здесь: Clinicaltrials.gov. В ЕС tDCS одобрен для лечения боли и депрессии. Вы можете использовать бесплатный инструмент ниже, чтобы найти врачей и клиники, которые предоставляют лечение на основе tDCS
Вы можете использовать бесплатный инструмент ниже, чтобы найти врачей и клиники, которые предоставляют лечение на основе tDCS
Одобрена ли tDCS FDA?
tDCS в настоящее время не одобрен FDA. Это означает, что FDA США не оценивало и не одобряло «маркетинговую» заявку от компании. Это не означает, что FDA США приняло официальное решение об эффективности или безопасности tDCS для индукции любой спецификации, такой как депрессия или боль. В США tDCS для медицинского применения считается «исследовательским». FDA не читает клинические испытания и принимает решения на основе литературы, FDA США отвечает только на «маркетинговые» запросы, сделанные конкретными компаниями. FDA обычно не регулирует немедицинское использование устройств, включая использование для «здоровья». цели 11 . В этом смысле важно отметить, что tDCS широко рассматривается исследователями и экспертами как заболевание с низким уровнем риска. Фактически, FDA предоставило письма «513g» нескольким компаниям, прямо разрешая им продавать tDCS для конкретных немедицинских целей.
FDA также не регулирует медицинскую практику, то есть не регулирует работу врачей. По этой причине многие врачи назначают лечение «не по прямому назначению» — то, что, по мнению врачей, работает, но не имеет «маркетингового» ярлыка от FDA для компании. tDCS одобрен для лечения в большинстве стран мира 12 , включая Европейский Союз 13 , Израиль и Сингапур. Таким образом, хотя tDCS в настоящее время не одобрена FDA, это не означает, что tDCS нельзя тестировать на законных основаниях или использовать в определенных контекстах.
Фактически, FDA предоставило письма «513g» нескольким компаниям, прямо разрешая им продавать tDCS для конкретных немедицинских целей.
FDA также не регулирует медицинскую практику, то есть не регулирует работу врачей. По этой причине многие врачи назначают лечение «не по прямому назначению» — то, что, по мнению врачей, работает, но не имеет «маркетингового» ярлыка от FDA для компании. tDCS одобрен для лечения в большинстве стран мира 12 , включая Европейский Союз 13 , Израиль и Сингапур. Таким образом, хотя tDCS в настоящее время не одобрена FDA, это не означает, что tDCS нельзя тестировать на законных основаниях или использовать в определенных контекстах.
Сколько стоит tDCS?
Стоимость устройств tDCS может варьироваться от примерно 100 долларов США за базовые «потребительские» устройства tDCS до тысяч долларов за системы tDCS исследовательского уровня. Существует широкий спектр функций и возможностей в различных
устройства.
Может ли tDCS лечить тревогу?
В ходе нескольких клинических испытаний 14 , проведенных в ведущем медицинском центре, сообщалось об эффектах tDCS по снижению тревожности. В США tDCS не одобрен для лечения медицинской тревоги. Методы, связанные с tDCS, такие как транскраниальная стимуляция переменным током (tACS), показали многообещающие результаты в клинических испытаниях 15 . Другой родственный метод, стимуляция краниальной электротерапии (CES), одобрен FDA для тревоги 16 .
Может ли tDCS лечить депрессию?
Несколько клинических испытаний показали, что tDCS может лечить депрессию 17 . tDCS также имеет гораздо меньше побочных эффектов, чем лекарства 18 . В США tDCS не одобрен для лечения депрессии. В большинстве стран мира, в том числе в Европе, tDCS одобрен для лечения депрессии 19 .
Каковы преимущества tDCS?
tDCS используется для многих различных приложений, которые включают изменение мозга, чтобы влиять на то, как люди думают или чувствуют 20 .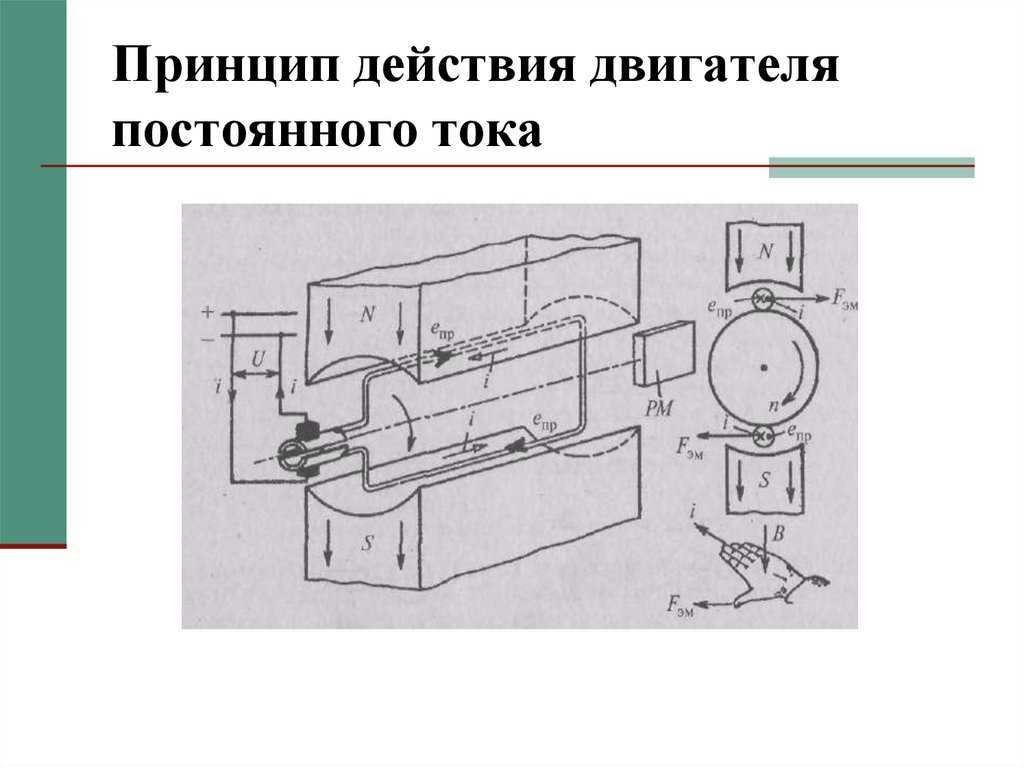 tDCS часто сочетается с какой-либо другой формой активности или обучения с целью tDCS повысить эту конкретную мозговую активность. Было показано, что tDCS заставляет людей учиться быстрее 21 . Например, tDCS может улучшить осознанность (электронная медитация). Люди также заинтересованы в tDCS для увеличения 22 «рабочей памяти».
tDCS часто сочетается с какой-либо другой формой активности или обучения с целью tDCS повысить эту конкретную мозговую активность. Было показано, что tDCS заставляет людей учиться быстрее 21 . Например, tDCS может улучшить осознанность (электронная медитация). Люди также заинтересованы в tDCS для увеличения 22 «рабочей памяти».
Поделиться |
Содержимое не предназначено для замены профессиональной медицинской консультации, диагностики или лечения. Всегда обращайтесь за советом к своему врачу или другому квалифицированному поставщику медицинских услуг по любым вопросам, которые могут у вас возникнуть относительно состояния здоровья.
1. Вудс А.Дж., Антал А., Биксон М., Богджио П.С., Брунони А.Р., Сельник П.… Ницше М.А. Техническое руководство по tDCS и связанным с ними инструментам неинвазивной стимуляции мозга. Клиническая нейрофизиология. 2016;127(2):1031–1048.
2. Выступление доктора Майкла Ницше о физиологических основах tDCS на YouTube. 3. Кронберг Г., Рахман А., Шарма М., Биксон М. и Парра Л., 2020. Стимуляция постоянным током повышает пластичность Хебба в лабораторных условиях. Стимуляция мозга
4. Эхтиари Х., Таваколи Х., Аддолорато Г., Баекен С., Бончи А., Кампанелла С., Каштело-Бранко Л., Шале-Буджу Г., Кларк В.П., Клаус Э., Дэннон П.Н., Дель Феличе А., Ден Уйл Т., Дайана М., ди Джаннантонио М., Федота Дж. Р., Фитцджеральд П., Галлимберти Л., Гралль-Броннек М., Херреманс СК, Херрманн М.Дж., Джамиль А., Хедр Э., Куимцидис С., Козак К., Крупицкий Э., Ламм С., Лехнер В.В., Мадео Г., Мальмир Н. , Мартинотти Г., Макдональд В., Монтемитро К., Накамура-Паласиос Э.М., Насехи М., Ноэль Х, Носратабади М., Паулюс М., Петторрусо М., Прадхан Б., Прахарадж С.К., Рафферти Х., Салем Г., Салмерон Б.Дж., Соважет А., Шлютер Р.С. , Сержиу К., Шахбабаи А., Шеффер К., Спаньоло П.А., Стил В.Р., Юань Т-Ф, ван Донген Дж., Ван Ваес В., Венкатасубраманян Г., ВердехоГарсия А., Вервир И., Уэлш Дж., Уэсли М.Дж., Виткевиц К., Явари Ф., Зарриндаст М.
3. Кронберг Г., Рахман А., Шарма М., Биксон М. и Парра Л., 2020. Стимуляция постоянным током повышает пластичность Хебба в лабораторных условиях. Стимуляция мозга
4. Эхтиари Х., Таваколи Х., Аддолорато Г., Баекен С., Бончи А., Кампанелла С., Каштело-Бранко Л., Шале-Буджу Г., Кларк В.П., Клаус Э., Дэннон П.Н., Дель Феличе А., Ден Уйл Т., Дайана М., ди Джаннантонио М., Федота Дж. Р., Фитцджеральд П., Галлимберти Л., Гралль-Броннек М., Херреманс СК, Херрманн М.Дж., Джамиль А., Хедр Э., Куимцидис С., Козак К., Крупицкий Э., Ламм С., Лехнер В.В., Мадео Г., Мальмир Н. , Мартинотти Г., Макдональд В., Монтемитро К., Накамура-Паласиос Э.М., Насехи М., Ноэль Х, Носратабади М., Паулюс М., Петторрусо М., Прадхан Б., Прахарадж С.К., Рафферти Х., Салем Г., Салмерон Б.Дж., Соважет А., Шлютер Р.С. , Сержиу К., Шахбабаи А., Шеффер К., Спаньоло П.А., Стил В.Р., Юань Т-Ф, ван Донген Дж., Ван Ваес В., Венкатасубраманян Г., ВердехоГарсия А., Вервир И., Уэлш Дж., Уэсли М.Дж., Виткевиц К., Явари Ф., Зарриндаст М. Р. , Zawertailo L, Zhang X, Cha YH, George TP, Frohlich F, Goudriaan AE, Fecteau S, Daughters SB, Stein EA, Fregni F, Nitsche MA, Zangen A, Bikson M, Hanlon CA (2019). Транскраниальная электрическая и магнитная стимуляция (tES и TMS) для лечения зависимости: согласованный документ о нынешнем состоянии науки и пути вперед. Нейронаука и биоповеденческие обзоры. 2019. 104: 118-140
5. Коффман Б., Трамбо М., Флорес Р., Гарсия К., ван дер Мерве А., Вассерманн Э., Вайзенд М. и Кларк В., 2020. Влияние Tdcs на Производительность и обучение обнаружению целей: взаимодействие с характеристиками стимула и экспериментальным планом.
6. Электронная медитация: новый инструмент для древней техники на сайте Medical X Press Медицинского университета Южной Каролины.
7. Биксон М., Гроссман П., Томас С., Занноу А.Л., Цзян Дж., Аднан Т. и др. (2016). Безопасность транскраниальной стимуляции постоянным током: обновление, основанное на фактических данных, 2016 г. Стимуляция мозга. 9641–661. 10.1016/j.brs.
Р. , Zawertailo L, Zhang X, Cha YH, George TP, Frohlich F, Goudriaan AE, Fecteau S, Daughters SB, Stein EA, Fregni F, Nitsche MA, Zangen A, Bikson M, Hanlon CA (2019). Транскраниальная электрическая и магнитная стимуляция (tES и TMS) для лечения зависимости: согласованный документ о нынешнем состоянии науки и пути вперед. Нейронаука и биоповеденческие обзоры. 2019. 104: 118-140
5. Коффман Б., Трамбо М., Флорес Р., Гарсия К., ван дер Мерве А., Вассерманн Э., Вайзенд М. и Кларк В., 2020. Влияние Tdcs на Производительность и обучение обнаружению целей: взаимодействие с характеристиками стимула и экспериментальным планом.
6. Электронная медитация: новый инструмент для древней техники на сайте Medical X Press Медицинского университета Южной Каролины.
7. Биксон М., Гроссман П., Томас С., Занноу А.Л., Цзян Дж., Аднан Т. и др. (2016). Безопасность транскраниальной стимуляции постоянным током: обновление, основанное на фактических данных, 2016 г. Стимуляция мозга. 9641–661. 10.1016/j.brs. 2017.07.001
8. Брунони А.Р., Амадера Дж., Бербель Б., Волц М.С., Риццерио Б.Г. и Фрегни Ф. (2011). Систематический обзор отчетов и оценки побочных эффектов, связанных с транскраниальной стимуляцией постоянным током. Международный журнал нейропсихофармакологии, 14(8), 1133-1145.
9. Монаи Х., Окура М., Танака М., Оэ Ю., Конно А., Хираи Х., … и Хирасе Х. (2016). Визуализация кальция показывает участие глии в пластичности, индуцированной транскраниальной стимуляцией постоянным током, в мозге мышей. Связи с природой, 7.
10. Использование продаваемых лекарств, биологических препаратов и медицинских устройств «не по прямому назначению» и исследовательское использование — Руководство для институциональных наблюдательных советов и клинических исследователей fda.gov
11. Общее оздоровление: политика в отношении устройств с низким уровнем риска — руководство для промышленности и персонала Управления по санитарному надзору за качеством пищевых продуктов и медикаментов fda.ggOyR0iXCbMQv3Xipma34MD
12.
2017.07.001
8. Брунони А.Р., Амадера Дж., Бербель Б., Волц М.С., Риццерио Б.Г. и Фрегни Ф. (2011). Систематический обзор отчетов и оценки побочных эффектов, связанных с транскраниальной стимуляцией постоянным током. Международный журнал нейропсихофармакологии, 14(8), 1133-1145.
9. Монаи Х., Окура М., Танака М., Оэ Ю., Конно А., Хираи Х., … и Хирасе Х. (2016). Визуализация кальция показывает участие глии в пластичности, индуцированной транскраниальной стимуляцией постоянным током, в мозге мышей. Связи с природой, 7.
10. Использование продаваемых лекарств, биологических препаратов и медицинских устройств «не по прямому назначению» и исследовательское использование — Руководство для институциональных наблюдательных советов и клинических исследователей fda.gov
11. Общее оздоровление: политика в отношении устройств с низким уровнем риска — руководство для промышленности и персонала Управления по санитарному надзору за качеством пищевых продуктов и медикаментов fda.ggOyR0iXCbMQv3Xipma34MD
12. Френьи Ф., Ницше М.А., Лоо К.К., Брунони А.Р., Маранголо П., Лейте Дж., Карвалью С., Болоньини Н., Каумо В., Пайк Н.Дж., Симис М., Уеда К., Эхтиари Х., Луу П., Такер Д.М., Тайлер В.Дж., Brunelin J, Datta A, Juan CH, Venkatasubramanian G, Boggio PM, Bikson M. Нормативные аспекты клинического и исследовательского использования транскраниальной стимуляции постоянным током (tDCS): обзор и рекомендации группы экспертов. Clin Res Regul Aff. 2015 1 марта; 32(1):22-35.
13. Soterix Medical, Inc. получает одобрение CE Mark для терапии депрессии 1×1 tDCS | soterixmedical.com
14. Нисида К., Косикава Ю., Моришима Ю. и др. Предстимульная активность мозга связана с изменением состояния тревожности при однократной транскраниальной стимуляции постоянным током. Передний шум нейронов. 2019;13:266. doi:10.3389/fnhum.2019.00266
15. Клэнси К.Дж. и др. Длительное увеличение связи и снижение беспокойства за счет транскраниальной стимуляции переменным током. соц. Познан. Оказывать воздействие. Неврологи.
Френьи Ф., Ницше М.А., Лоо К.К., Брунони А.Р., Маранголо П., Лейте Дж., Карвалью С., Болоньини Н., Каумо В., Пайк Н.Дж., Симис М., Уеда К., Эхтиари Х., Луу П., Такер Д.М., Тайлер В.Дж., Brunelin J, Datta A, Juan CH, Venkatasubramanian G, Boggio PM, Bikson M. Нормативные аспекты клинического и исследовательского использования транскраниальной стимуляции постоянным током (tDCS): обзор и рекомендации группы экспертов. Clin Res Regul Aff. 2015 1 марта; 32(1):22-35.
13. Soterix Medical, Inc. получает одобрение CE Mark для терапии депрессии 1×1 tDCS | soterixmedical.com
14. Нисида К., Косикава Ю., Моришима Ю. и др. Предстимульная активность мозга связана с изменением состояния тревожности при однократной транскраниальной стимуляции постоянным током. Передний шум нейронов. 2019;13:266. doi:10.3389/fnhum.2019.00266
15. Клэнси К.Дж. и др. Длительное увеличение связи и снижение беспокойства за счет транскраниальной стимуляции переменным током. соц. Познан. Оказывать воздействие. Неврологи. 2018;13:1305–1316.
16. Неврологические устройства; Реклассификация краниальных электротерапевтических стимуляторов, предназначенных для лечения тревоги и/или бессонницы; Дата вступления в силу требования о предварительном одобрении краниальных электротерапевтических стимуляторов, предназначенных для лечения депрессии | Federalregister.gov
17. Коффман Б., Трамбо М., Флорес Р., Гарсия К., ван дер Мерве А., Вассерманн Э., Вайзенд М. и Кларк В., 2020. Влияние Tdcs на Производительность и обучение обнаружению целей: взаимодействие с характеристиками стимула и экспериментальным планом.
18. Андре Р. Брунони, доктор медицины, доктор философии, Адриано Х. Моффа, психолог, Бернардо Сампайо-Джуниор, доктор медицины, Лукас Боррионе, доктор медицины, Марина Л. Морено, психолог, Ракель А. Фернандес , Psy.D., Беатрис П. Веронези, Psy.D., Барбара С. Ногейра, Psy.D., Луана В.М. Апарисио, доктор медицины, Лаис Б. Разза, психолог, Ренан Чаморро, психолог, Луара С. Торт. Испытание электрической терапии постоянным током в сравнении с эсциталопрамом при депрессии
Список авторов.
2018;13:1305–1316.
16. Неврологические устройства; Реклассификация краниальных электротерапевтических стимуляторов, предназначенных для лечения тревоги и/или бессонницы; Дата вступления в силу требования о предварительном одобрении краниальных электротерапевтических стимуляторов, предназначенных для лечения депрессии | Federalregister.gov
17. Коффман Б., Трамбо М., Флорес Р., Гарсия К., ван дер Мерве А., Вассерманн Э., Вайзенд М. и Кларк В., 2020. Влияние Tdcs на Производительность и обучение обнаружению целей: взаимодействие с характеристиками стимула и экспериментальным планом.
18. Андре Р. Брунони, доктор медицины, доктор философии, Адриано Х. Моффа, психолог, Бернардо Сампайо-Джуниор, доктор медицины, Лукас Боррионе, доктор медицины, Марина Л. Морено, психолог, Ракель А. Фернандес , Psy.D., Беатрис П. Веронези, Psy.D., Барбара С. Ногейра, Psy.D., Луана В.М. Апарисио, доктор медицины, Лаис Б. Разза, психолог, Ренан Чаморро, психолог, Луара С. Торт. Испытание электрической терапии постоянным током в сравнении с эсциталопрамом при депрессии
Список авторов.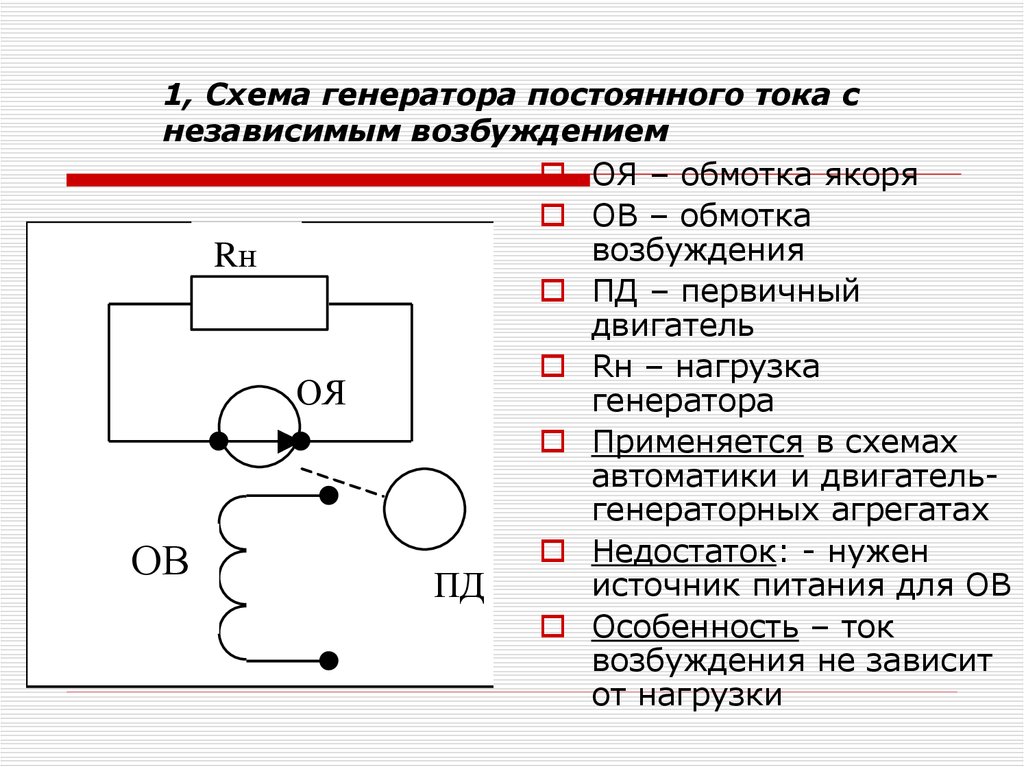 19. Soterix Medical получает знак CE за систему терапии депрессии | fdanews.com
20. Инструмент для разума | Маром Биксон | TEDxBushwick на YouTube
21. Доклад Винса Кларка о TDCS for Cognitive Enhancement на YouTube.
22. Трамбо, М., Матцен, Л., Коффман, Б., Хантер, М., Джонс, А., Робинсон, К. и Кларк, В., 2020. Повышение производительности рабочей памяти с помощью транскраниальной стимуляции постоянным током: Возможность ближнего и дальнего перевода.
23. Вудс А.Дж., Антал А., Биксон М., Боггио П.С., Брунони А.Р., Целник П., Коэн Л.Г., Фрегни Ф., Херрманн К.С., Каппенман Э.С., Кноткова Х., Либетанц Д., Миниусси К., Миранда П.С., Паулюс В., Приори А., Реато Д., Стэгг С., Вендерот Н., Ницше М.А. Техническое руководство по tDCS и связанным с ними инструментам неинвазивной стимуляции мозга. Клин Нейрофизиол. 2016 Февраль; 127 (2): 1031-48
19. Soterix Medical получает знак CE за систему терапии депрессии | fdanews.com
20. Инструмент для разума | Маром Биксон | TEDxBushwick на YouTube
21. Доклад Винса Кларка о TDCS for Cognitive Enhancement на YouTube.
22. Трамбо, М., Матцен, Л., Коффман, Б., Хантер, М., Джонс, А., Робинсон, К. и Кларк, В., 2020. Повышение производительности рабочей памяти с помощью транскраниальной стимуляции постоянным током: Возможность ближнего и дальнего перевода.
23. Вудс А.Дж., Антал А., Биксон М., Боггио П.С., Брунони А.Р., Целник П., Коэн Л.Г., Фрегни Ф., Херрманн К.С., Каппенман Э.С., Кноткова Х., Либетанц Д., Миниусси К., Миранда П.С., Паулюс В., Приори А., Реато Д., Стэгг С., Вендерот Н., Ницше М.А. Техническое руководство по tDCS и связанным с ними инструментам неинвазивной стимуляции мозга. Клин Нейрофизиол. 2016 Февраль; 127 (2): 1031-48
Транскраниальная стимуляция постоянным током: дорожная карта для исследований, от механизма действия до клинического применения
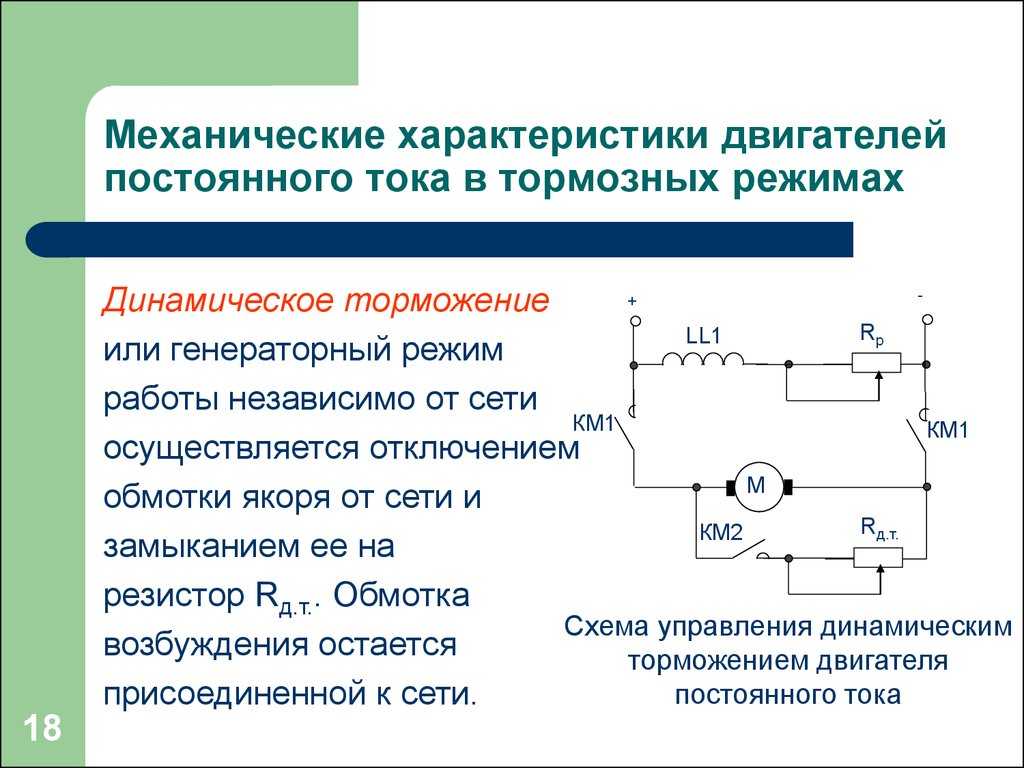 «>
«>Явари Ф., Джамиль А., Мосайеби Самани М., Видор Л.П., Ницше М.А. Основные и функциональные эффекты транскраниальной электростимуляции (ТЭС) — введение. Neurosci Biobehav Rev. 2018; 85:81–92.
Артикул пабмед Google ученый
Lefaucheur JP, Antal A, Ayache SS, Benninger DH, Brunelin J, Cogiamanian F, et al. Основанные на фактических данных рекомендации по терапевтическому использованию транскраниальной стимуляции постоянным током (tDCS). Клин Нейрофизиол. 2017; 128:56–92.
Артикул пабмед Google ученый
Mutz J, Edgcumbe DR, Brunoni AR, Fu CHY. Эффективность и приемлемость неинвазивной стимуляции мозга для лечения униполярной и биполярной депрессии у взрослых: систематический обзор и метаанализ рандомизированных плацебо-контролируемых исследований. Neurosci Biobehav Rev. 2018; 92: 291–303.
Артикул пабмед Google ученый
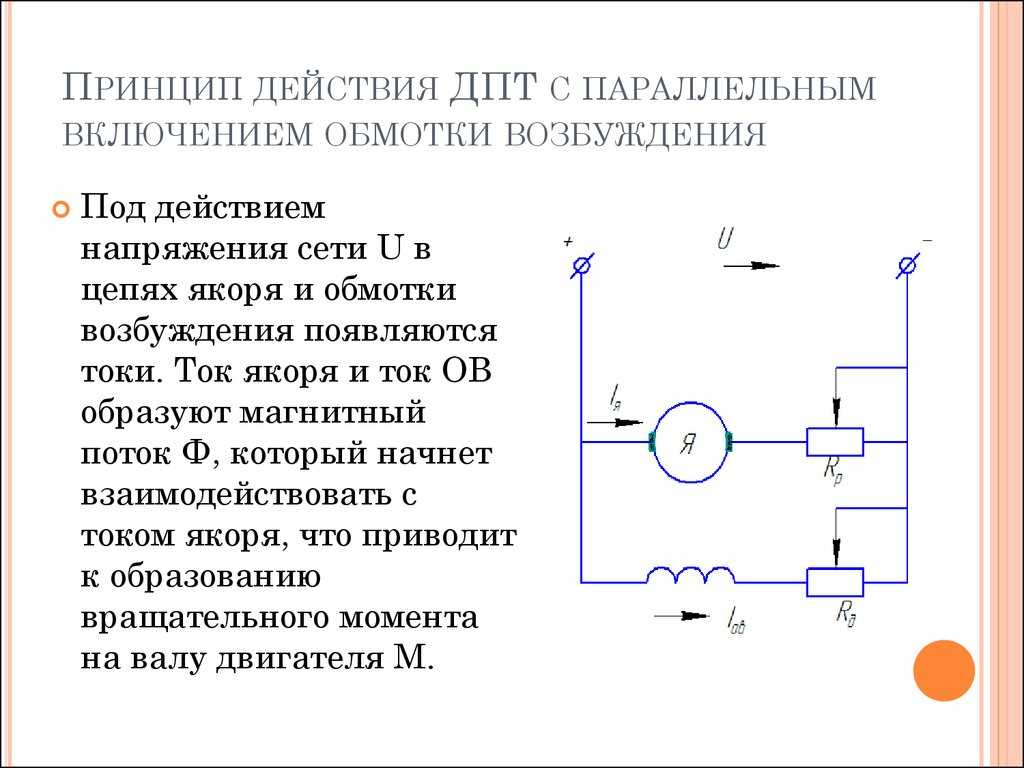 «>
«>Моффа А.Х., Брунони А.Р., Фрегни Ф., Палм Ю., Падберг Ф., Блумбергер Д.М. и др. Безопасность и приемлемость транскраниальной стимуляции постоянным током для неотложного лечения больших депрессивных эпизодов: анализ индивидуальных данных пациентов. J Аффективное расстройство. 2017; 221:1–5.
Артикул пабмед Google ученый
Апарисио Л.В.М., Гуариенти Ф., Разза Л.Б., Карвалью А.Ф., Френьи Ф., Брунони А.Р. Систематический обзор приемлемости и переносимости транскраниальной стимуляции постоянным током в нейропсихиатрических исследованиях. Мозговой стимул. 2016;9: 671–81.
Артикул пабмед Google ученый
Фергюсон Дж.М. Антидепрессанты из группы СИОЗС: побочные эффекты и переносимость. Prim Care Companion J Clin Psychiatry. 2001; 3: 22–7.
Артикул пабмед ПабМед Центральный Google ученый
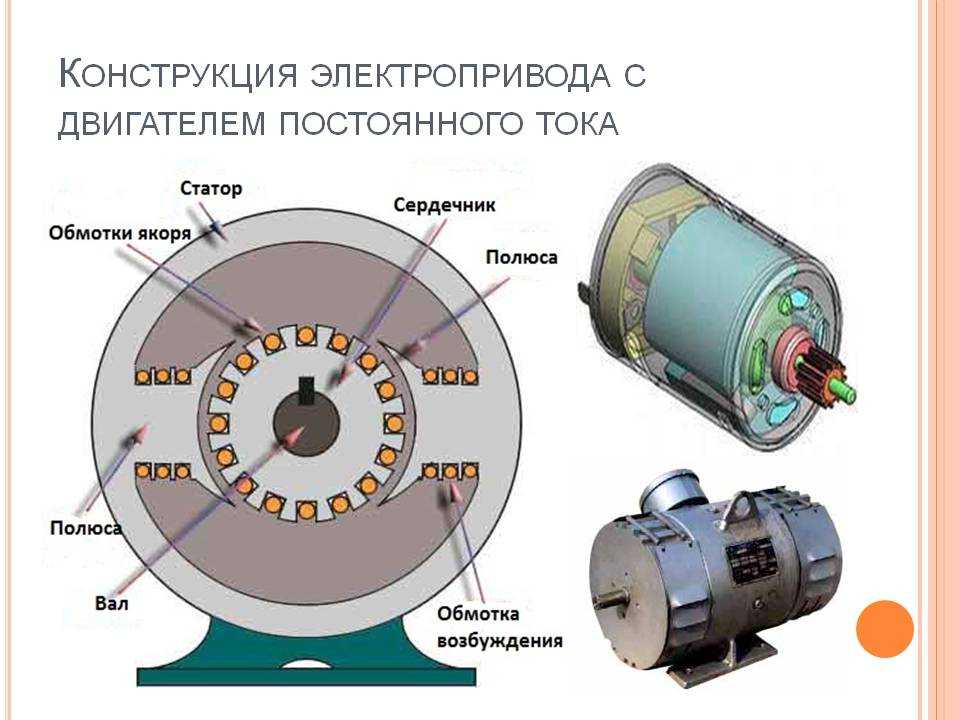 «>
«>Брунони А.Р., Ницше М.А., Болоньини Н., Биксон М., Вагнер Т., Мерабет Л. и др. Клинические исследования с транскраниальной стимуляцией постоянным током (tDCS): проблемы и будущие направления. Мозговой стимул. 2012;5:175–1795.
Артикул пабмед Google ученый
Esmaeilpour Z, Marangolo P, Hampstead BM, Bestmann S, Galletta E, Knotkova H, et al. Неполные доказательства того, что увеличение силы тока tDCS улучшает результаты. Мозговой стимул. 2018;11:310–21.
Артикул пабмед Google ученый
Ворослакос М., Такеучи Ю., Бринички К., Зомбори Т., Олива А., Фернандес-Руис А. и др. Прямое влияние транскраниальной электрической стимуляции на мозговые цепи у крыс и людей. Нац коммун. 2018;9:483.
Артикул КАС пабмед ПабМед Центральный Google ученый
 «>
«>Хуанг Ю., Лю А.А., Лафон Б., Фридман Д., Даян М., Ван Х и др. Измерения и модели электрических полей в мозге человека in vivo при транскраниальной электростимуляции. Элиф. 2017; 6:e18834.
Opitz A, Falchier A, Yan CG, Yeagle EM, Linn GS, Megevand P, et al. Пространственно-временная структура внутричерепных электрических полей, индуцированных транскраниальной электрической стимуляцией у человека и нечеловеческих приматов. Научный доклад 2016; 6: 31236.
Артикул КАС пабмед ПабМед Центральный Google ученый
Джексон М.П., Рахман А., Лафон Б., Кронберг Г., Линг Д., Парра Л.С. и др. Животные модели транскраниальной стимуляции постоянным током: методы и механизмы. Клин Нейрофизиол. 2016; 127:3425–54.
Артикул пабмед ПабМед Центральный Google ученый
Реато Д.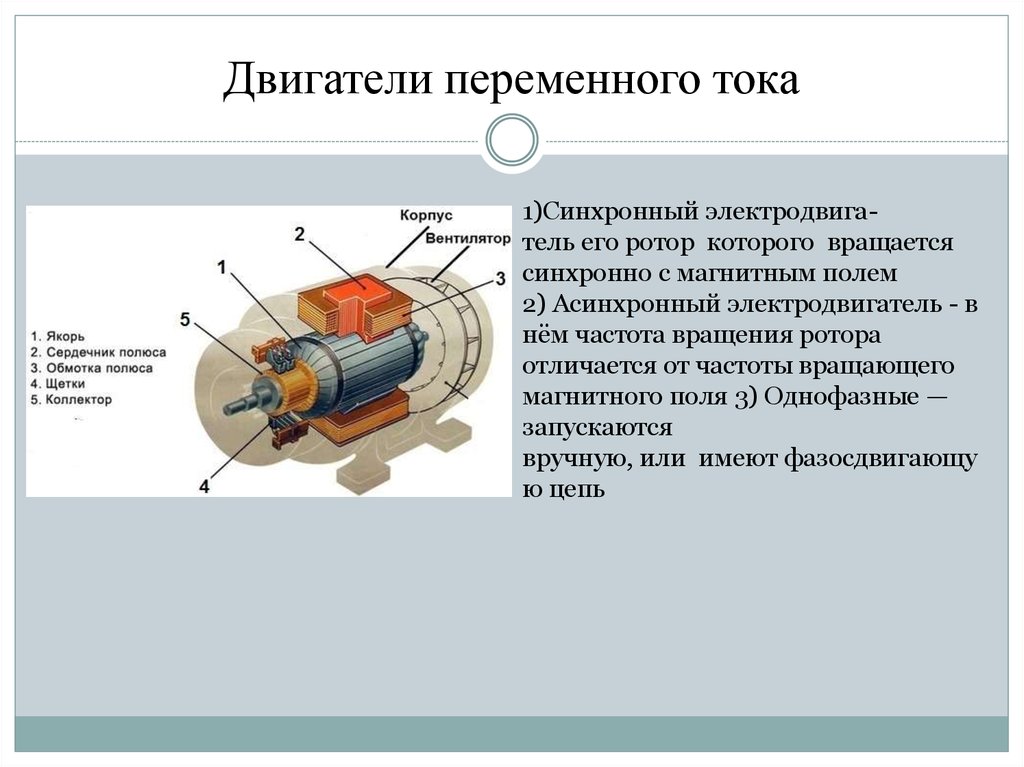 , Рахман А., Биксон М., Парра Л.С. Электрическая стимуляция низкой интенсивности влияет на динамику сети, модулируя скорость популяции и время спайков. Дж. Нейроски. 2010; 30:15067–79.
, Рахман А., Биксон М., Парра Л.С. Электрическая стимуляция низкой интенсивности влияет на динамику сети, модулируя скорость популяции и время спайков. Дж. Нейроски. 2010; 30:15067–79.
Артикул КАС пабмед ПабМед Центральный Google ученый
Лю А., Ворослакос М., Кронберг Г., Хенин С., Краузе М.Р., Хуанг Ю. и др. Непосредственные нейрофизиологические эффекты транскраниальной электростимуляции. Нац коммун. 2018;9:5092.
Артикул КАС пабмед ПабМед Центральный Google ученый
McDonnell MD, Abbott D. Что такое стохастический резонанс? Определения, заблуждения, дебаты и их отношение к биологии. PLoS Comput Biol. 2009 г.;5:e1000348.
Артикул КАС пабмед ПабМед Центральный Google ученый
Кронберг Г., Бриди М. , Абель Т., Биксон М., Парра Л.С. Стимуляция постоянным током модулирует LTP и LTD: зависимость от активности и дендритные эффекты. Мозговой стимул. 2017;10:51–8.
, Абель Т., Биксон М., Парра Л.С. Стимуляция постоянным током модулирует LTP и LTD: зависимость от активности и дендритные эффекты. Мозговой стимул. 2017;10:51–8.
Артикул пабмед Google ученый
Биксон М., Имя А., Рахман А. Происхождение специфичности во время tDCS: анатомические, избирательные по активности и входные механизмы смещения. Передний шум нейронов. 2013;7:688.
Артикул пабмед ПабМед Центральный Google ученый
Онтон Дж., Делорм А., Макейг С. Динамика фронтальной срединной ЭЭГ во время рабочей памяти. Нейроизображение. 2005; 27: 341–56.
Артикул пабмед Google ученый
Симонсмейер Б.А., Грабнер Р.Х., Хайн Дж., Кренц У., Шнайдер М. Электрическая стимуляция мозга (ЭС) улучшает обучение больше, чем производительность: метаанализ. Neurosci Biobehav Rev. 2018; 84: 171–81.
Neurosci Biobehav Rev. 2018; 84: 171–81.
Артикул пабмед Google ученый
Коэн МХ. Откуда берется ЭЭГ и что это значит? Тренды Нейроси. 2017;40:208–18.
Артикул КАС пабмед Google ученый
Логотетис Н.К., Ванделл Б.А. Интерпретация ЖИРНОГО сигнала. Annu Rev Physiol. 2004; 66: 735–69.
Артикул КАС пабмед Google ученый
Рауджи В., Чокка М., Захария А., Соарес Д., Труонг Д., Биксон М. и др. Изменения двигательной возбудимости при tDCS специфичны для направления тока. Мозговой стимул. 2018;11:289–98.
Артикул пабмед ПабМед Центральный Google ученый
Алексейчук И., Диерс К., Паулюс В., Антал А. Транскраниальная электрическая стимуляция затылочной коры во время зрительного восприятия изменяет величину активности BOLD: комбинированный подход tES-fMRI. Нейроизображение. 2016; 140:110–7.
Нейроизображение. 2016; 140:110–7.
Артикул пабмед Google ученый
Fiori V, Kunz L, Kuhnke P, Marangolo P, Hartwigsen G. Транскраниальная стимуляция постоянным током (tDCS) облегчает изучение глаголов, изменяя эффективную связь в здоровом мозге. Нейроизображение. 2018;181:550–9.
Артикул пабмед Google ученый
Вебер М.Дж., Мессинг С.Б., Рао Х., Детре Дж.А., Томпсон-Шилль С.Л. Префронтальная транскраниальная стимуляция постоянным током изменяет активацию и связь в корковых и подкорковых системах вознаграждения: исследование tDCS-fMRI. Hum Brain Map. 2014; 35:3673–86.
Артикул пабмед ПабМед Центральный Google ученый
Бунстра Т.В., Николин С., Мейзенер А.С., Мартин Д.М., Лоо К.К. Изменение средней частоты электроэнцефалографии в состоянии покоя после транскраниальной стимуляции постоянным током.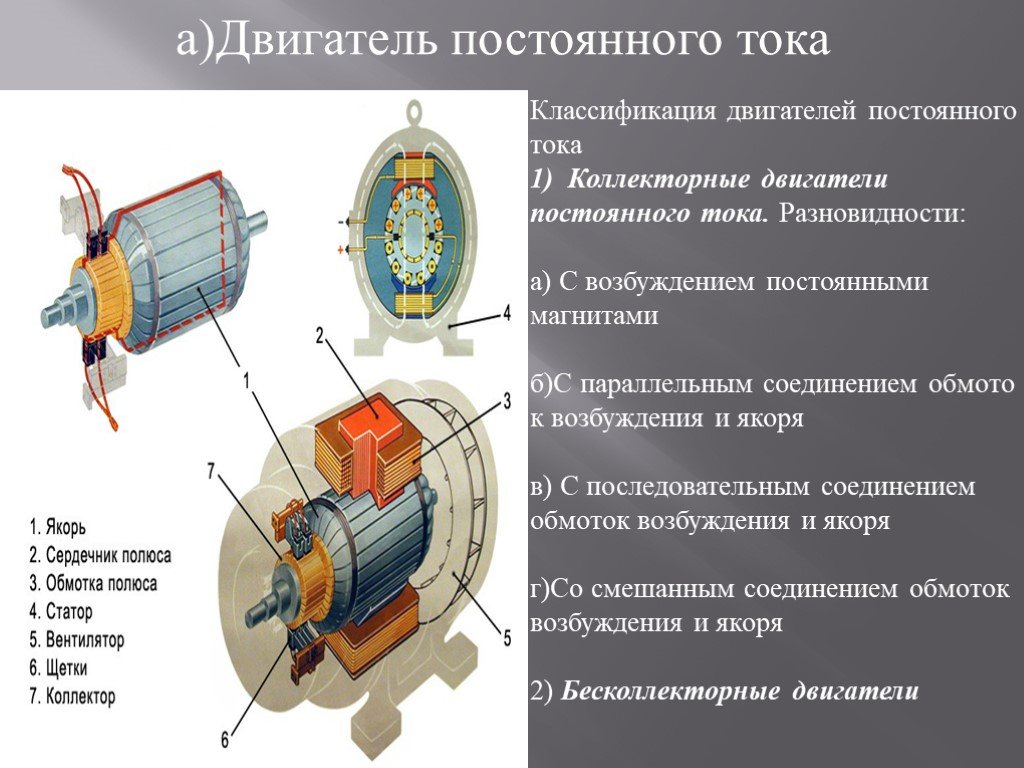 Передний шум нейронов. 2016;10:270.
Передний шум нейронов. 2016;10:270.
ПабМед ПабМед Центральный Google ученый
Wilson TW, McDermott TJ, Mills MS, Coolidge NM, Heinrichs-Graham E. tDCS модулирует визуальные гамма-колебания и базальную альфа-активность в затылочной коре: данные MEG. Кора головного мозга. 2018;28:1597–609.
Артикул пабмед Google ученый
Хэнли СиДжей, Сингх К.Д., МакГонигл ДиДжей. Транскраниальная модуляция колебательных ответов мозга: параллельное исследование tDCS-MEG. Нейроизображение. 2016;140:20–32.
Артикул пабмед Google ученый
Маршалл Т.Р., Эстерер С., Херринг Дж.Д., Бергманн Т.О., Дженсен О. О взаимосвязи между возбудимостью коры и зрительными колебательными реакциями — параллельное исследование tDCS-MEG. Нейроизображение. 2016; 140:41–9.
Артикул пабмед Google ученый
Рейнхарт Р.М., Чжу Дж., Парк С., Вудман Г.Ф. Синхронизация тета-колебаний со стимуляцией постоянным током усиливает адаптивный контроль в человеческом мозгу. Proc Natl Acad Sci USA. 2015; 112:9448–53.
Артикул КАС пабмед Google ученый
McDermott TJ, Wiesman AI, Mills MS, Spooner RK, Coolidge NM, Proskovec AL, et al. tDCS модулирует поведенческие характеристики и динамику нейронных колебаний, обслуживая зрительное избирательное внимание. Hum Brain Map. 2019;40:729–40.
Артикул пабмед Google ученый
Choe J, Coffman BA, Bergstedt DT, Ziegler MD, Phillips ME. Транскраниальная стимуляция постоянным током модулирует нейронную активность и обучение при обучении пилотов. Передний шум нейронов. 2016;10:34.
2016;10:34.
Артикул пабмед ПабМед Центральный Google ученый
Будевин М., Робертс Б.М., Мизрак Э., Ранганат С., Картер С.С. Префронтальная транскраниальная стимуляция постоянным током (tDCS) усиливает поведенческие и ЭЭГ-маркеры проактивного контроля. Cogn Neurosci. 2018;10:57–65.
Фрайс П. Механизм когнитивной динамики: нейронная связь через нейронную когерентность. Тенденции Cogn Sci. 2005; 9: 474–80.
Артикул пабмед Google ученый
Сигел М., Доннер Т.Х., Энгель А.К. Спектральные отпечатки крупномасштабных нейронных взаимодействий. Нат Рев Нейроски. 2012;13:121–34.
Артикул КАС пабмед Google ученый
Хьяфил А., Жиро А.Л., Фонтолан Л., Гуткин Б. Нейронная кросс-частотная связь: соединяющие архитектуры, механизмы и функции. Тренды Нейроси. 2015;38:725–40.
Тренды Нейроси. 2015;38:725–40.
Артикул КАС пабмед Google ученый
Бужаки Г., Шомбург Э.В. Что гамма-когерентность говорит нам о межрегиональной нейронной коммуникации? Нат Нейроски. 2015;18:484–9.
Артикул КАС пабмед ПабМед Центральный Google ученый
Fischer DB, Fried PJ, Ruffini G, Ripolles O, Salvador R, Banus J, et al. Мультифокальная tDCS, нацеленная на двигательную сеть в состоянии покоя, повышает возбудимость коры по сравнению с традиционной tDCS, нацеленной на одностороннюю моторную кору. Нейроизображение. 2017; 157:34–44.
Артикул КАС пабмед Google ученый
Имбургио М.Дж., Орр Дж.М. Влияние префронтальной tDCS на исполнительную функцию: методологические соображения, выявленные в результате метаанализа. Нейропсихология. 2018;117:156–66.
Нейропсихология. 2018;117:156–66.
Артикул пабмед Google ученый
Рейнхарт Р.М., Косман Дж.Д., Фукуда К., Вудман Г.Ф. Использование транскраниальной стимуляции постоянным током (tDCS) для понимания когнитивной обработки. Atten Percept Psychophys. 2017;79:3–23.
Артикул пабмед ПабМед Центральный Google ученый
Дмоховски Дж.П., Датта А., Биксон М., Су Ю., Парра Л.С. Оптимизированная мультиэлектродная стимуляция увеличивает фокус и интенсивность воздействия на цель. Дж. Нейронная инженерия. 2011;8:046011.
Артикул пабмед Google ученый
Battaglini L, Noventa S, Casco C. Анодальная и катодная электрическая стимуляция над V5 улучшает восприятие движения за счет усиления сигнала и снижения шума. Мозговой стимул. 2017;10:773–9.
Артикул пабмед Google ученый
Zito GA, Senti T, Cazzoli D, Muri RM, Mosimann UP, Nyffeler T, et al. Катодная HD-tDCS на правом V5 улучшает восприятие движения у людей. Фронт Behav Neurosci. 2015;9:257.
Артикул пабмед ПабМед Центральный Google ученый
Шуц Д., Иоаннидис Дж.П. Эмпирическая оценка опубликованных размеров и силы эффекта в недавней литературе по когнитивной нейробиологии и психологии. PLoS биол. 2017;15:e2000797.
Артикул КАС пабмед ПабМед Центральный Google ученый
Коллинг Л.Дж., Шукс Д. Статистический вывод и кризис репликации. Преподобный Филос Психол. 2018 г. https://doi.org/10.1007/s13164-018-0421-4.
Медина Дж., Кейсон С. Отсутствие доказательной ценности в исследованиях познания и рабочей памяти образцов транскраниальной стимуляции постоянным током (tDCS) у здоровых людей. кора. 2017;94:131–41.
кора. 2017;94:131–41.
Артикул пабмед Google ученый
Horvath JC, Forte JD, Carter O. Количественный обзор не находит доказательств когнитивных эффектов у здоровых людей от однократной транскраниальной стимуляции постоянным током (tDCS). Мозговой стимул. 2015; 8: 535–50.
Артикул пабмед Google ученый
Мерон Д., Хеджер Н., Гарнер М., Болдуин Д.С. Транскраниальная стимуляция постоянным током (tDCS) при лечении депрессии: систематический обзор и метаанализ эффективности и переносимости. Neurosci Biobehav Rev. 2015; 57:46–62.
Артикул пабмед Google ученый
Нитше М.А., Биксон М., Бестманн С. об использовании метаанализа в нейромодуляторной неинвазивной стимуляции мозга. Мозговой стимул. 2015; 8: 666–7.
Артикул пабмед Google ученый
 «>
«>Сантарнекки Э., Брем А.К., Левенбаум Э., Томпсон Т., Кадош Р.К., Паскуаль-Леоне А. Улучшение познания с помощью транскраниальной электрической стимуляции. Curr Opin Behav Sci. 2015;4:171–8.
Артикул Google ученый
Filmer HL, Dux PE, Mattingley JB. Применение транскраниальной стимуляции постоянным током для понимания функции мозга. Тренды Нейроси. 2014; 37: 742–53.
Артикул КАС пабмед Google ученый
Filmer HL, Varghese E, Hawkins GE, Mattingley JB, Dux PE. Улучшение внимания и принятия решений после комбинированной поведенческой тренировки и стимуляции мозга. Кора головного мозга. 2017;27:3675–82.
ПабМед Google ученый
Smith RC, Boules S, Mattiuz S, Youssef M, Tobe RH, Sershen H, et al. Влияние транскраниальной стимуляции постоянным током (tDCS) на познание, симптомы и курение при шизофрении: рандомизированное контролируемое исследование.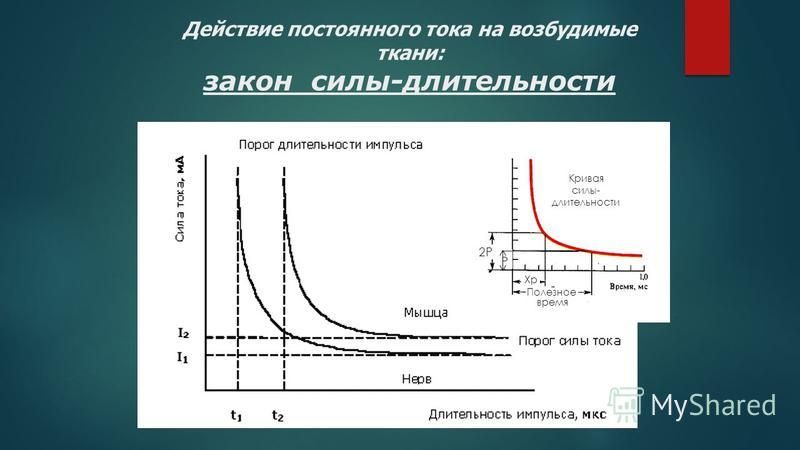 Шизофр Рез. 2015;168:260–6.
Шизофр Рез. 2015;168:260–6.
Артикул пабмед Google ученый
Хой К.Е., Бейли Н.В., Арнольд С.Л., Фицджеральд П.Б. Влияние транскраниальной стимуляции постоянным током на гамма-активность и рабочую память при шизофрении. Психиатрия рез. 2015; 228:191–6.
Артикул пабмед Google ученый
Vercammen A, Rushby JA, Loo C, Short B, Weickert CS, Weickert TW. Транскраниальная стимуляция постоянным током влияет на вероятностное ассоциативное обучение при шизофрении. Шизофр Рез. 2011;131:198–205.
Артикул пабмед Google ученый
Фридман Н.П., Мияке А., Корли Р.П., Янг С.Е., Дефрис Дж.К., Хьюитт Дж.К. Не все исполнительные функции связаны с интеллектом. Психологические науки. 2006; 17: 172–9.
Артикул пабмед Google ученый
 «>
«>Barch DM, Moore H, Nee DE, Manoach DS, Luck SJ. Выбор биомаркеров визуализации CNTRICS: рабочая память. Шизофр Булл. 2012; 38:43–52.
Артикул пабмед Google ученый
Манкузо Л.Е., Илиева И.П., Гамильтон Р.Х., Фарах М.Дж. Улучшает ли транскраниальная стимуляция постоянным током здоровую рабочую память?: метааналитический обзор. J Cogn Neurosci. 2016;28:1063–89.
Артикул пабмед Google ученый
Рейнхарт РМГ. Нарушение и спасение межпространственной тета-фазовой связи и адаптивного поведения. Proc Natl Acad Sci USA. 2017; 114:11542–7.
Артикул КАС пабмед Google ученый
Jacobson L, Javitt DC, Lavidor M. Активация торможения: уменьшение импульсивного поведения путем стимуляции постоянным током нижней лобной извилины. J Cogn Neurosci. 2011;23:3380–7.
J Cogn Neurosci. 2011;23:3380–7.
Артикул пабмед Google ученый
Li LM, Violante IR, Leech R, Hampshire A, Opitz A, McArthur D, et al. Когнитивное улучшение с помощью электрической стимуляции значимой сети зависит от структурной связности сети. Нейроизображение. 2019;185:425–33.
Артикул пабмед ПабМед Центральный Google ученый
Вергаллито А., Рива П., Пизони А., Ромеро Лауро Л.Дж. Модуляция негативных эмоций через анодную tDCS в правой вентролатеральной префронтальной коре. Нейропсихология. 2018;119:128–35.
Артикул пабмед Google ученый
Ironside M, Browning M, Ansari TL, Harvey CJ, Sekyi-Djan MN, Bishop SJ et al. Влияние стимуляции префронтальной коры на регуляцию реакции миндалевидного тела на угрозу у лиц с личностной тревожностью: рандомизированное клиническое исследование.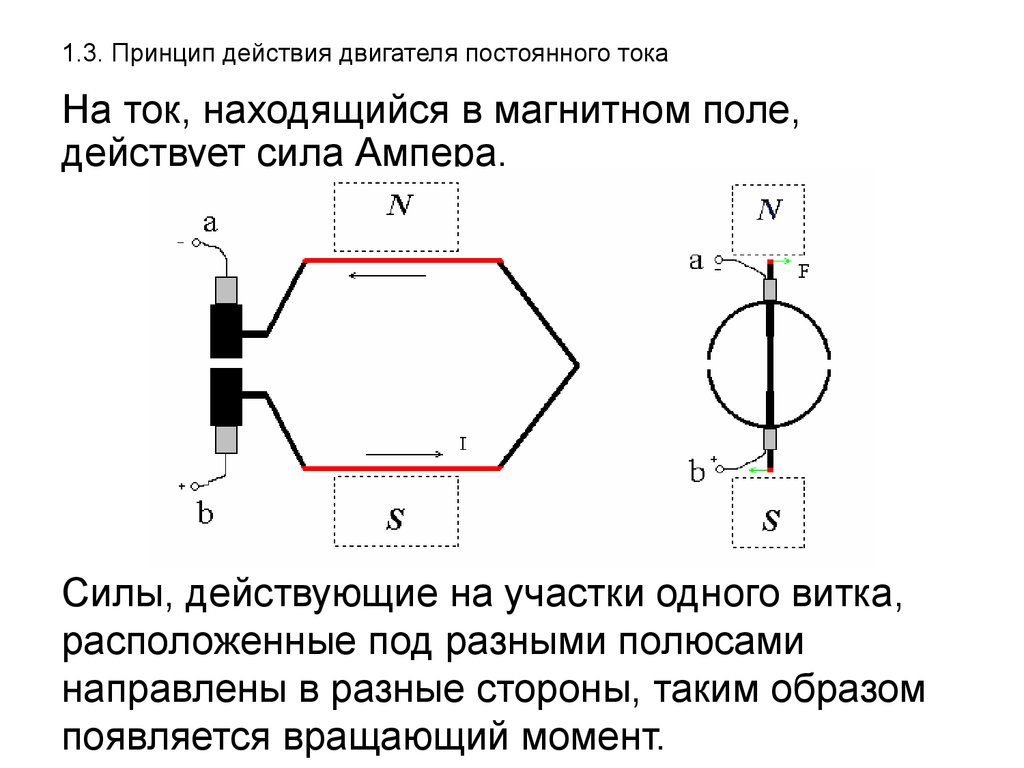 Джама Психиатрия. 2018;76:71–78.
Джама Психиатрия. 2018;76:71–78.
Brevet-Aeby C, Brunelin J, Iceta S, Padovan C, Poulet E. Префронтальная кора и импульсивность: интерес к неинвазивной стимуляции мозга. Neurosci Biobehav Rev. 2016; 71:112–34.
Артикул пабмед Google ученый
Лундквист М., Герман П., Ланснер А. Тета- и гамма-мощность увеличивается, а альфа/бета-мощность уменьшается с нагрузкой на память в модели аттракторной сети. J Cogn Neurosci. 2011;23:3008–20.
Артикул пабмед Google ученый
Compte A, Brunel N, Goldman-Rakic PS, Wang XJ. Синаптические механизмы и сетевая динамика, лежащие в основе пространственной рабочей памяти в модели корковой сети. Кора головного мозга. 2000;10:910–23.
Артикул КАС пабмед Google ученый
 «>
«>Хартвигсен Г. Гибкое перераспределение в когнитивных сетях. Тенденции Cogn Sci. 2018; 22: 687–98.
Артикул пабмед Google ученый
Брунони А.Р., Моффа А.Х., Фрегни Ф., Палм Ю., Падберг Ф., Блумбергер Д.М. и др. Транскраниальная стимуляция постоянным током при острых больших депрессивных эпизодах: метаанализ данных отдельных пациентов. Бр Дж. Психиатрия. 2016; 208: 522–31.
Артикул пабмед ПабМед Центральный Google ученый
Shiozawa P, Fregni F, Bensenor IM, Lotufo PA, Berlim MT, Daskalakis JZ, et al. Транскраниальная стимуляция постоянным током при большой депрессии: обновленный систематический обзор и метаанализ. Int J Neuropsychopharmacol. 2014;17:1443–52.
Артикул пабмед Google ученый
Брунони А.Р., Моффа А. Х., Сампайо-Джуниор Б., Боррионе Л., Морено М.Л., Фернандес Р.А. и др. Испытание электрической терапии постоянным током в сравнении с эсциталопрамом при депрессии. N Engl J Med. 2017; 376: 2523–33.
Х., Сампайо-Джуниор Б., Боррионе Л., Морено М.Л., Фернандес Р.А. и др. Испытание электрической терапии постоянным током в сравнении с эсциталопрамом при депрессии. N Engl J Med. 2017; 376: 2523–33.
Артикул КАС пабмед Google ученый
Брунони А.Р., Валиенго Л., Баккаро А., Занао Т.А., де Оливейра Дж.Ф., Гуларт А. и др. Сертралин по сравнению с терапией электрическим током для лечения депрессии: клиническое исследование: результаты факторного, рандомизированного, контролируемого исследования. Джама Психиатрия. 2013; 70: 383–91.
Артикул КАС пабмед Google ученый
Loo CK, Husain MM, McDonald WM, Aaronson S, O’Reardon JP, Alonzo A, et al. Международное рандомизированное контролируемое исследование транскраниальной стимуляции постоянным током при депрессии. Мозговой стимул. 2018;11:125–33.
Артикул пабмед Google ученый
 «>
«>Rock PL, Roiser JP, Riedel WJ, Blackwell AD. Когнитивные нарушения при депрессии: систематический обзор и метаанализ. Психомед. 2014;44:2029–40.
Артикул КАС пабмед Google ученый
Martin DM, Teng JZ, Lo TY, Alonzo A, Goh T, Iacoviello BM и др. Клиническое пилотное исследование транскраниальной стимуляции постоянным током в сочетании с когнитивно-эмоциональной тренировкой при лекарственно-резистентной депрессии. J Аффективное расстройство. 2018; 232:89–95.
Артикул КАС пабмед Google ученый
Wolkenstein L, Plewnia C. Улучшение когнитивного контроля при депрессии с помощью транскраниальной стимуляции постоянным током. Биол психиатрия. 2013;73:646–51.
Артикул пабмед Google ученый
Брунони А.Р., Боджио П.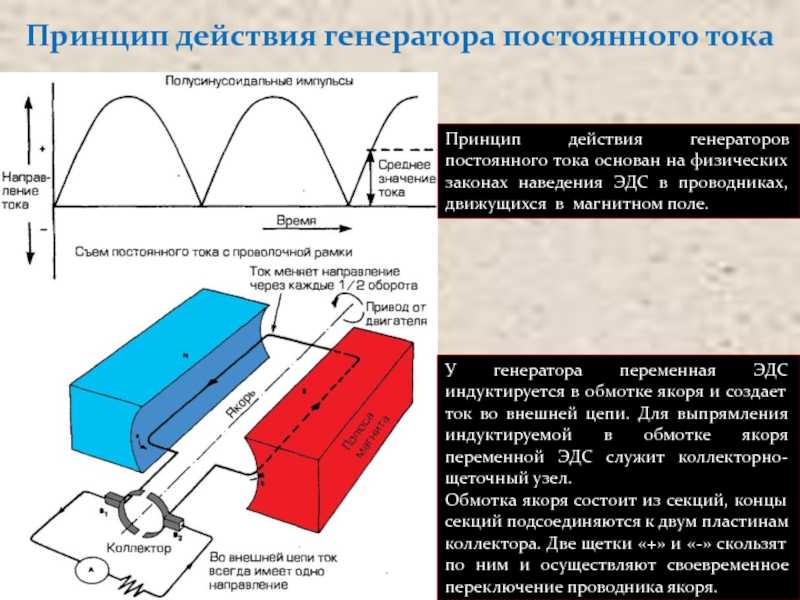 С., Де Рэдт Р., Бенсенор И.М., Лотуфо П.А., Намюр В. и др. Терапия когнитивного контроля и транскраниальная стимуляция постоянным током при депрессии: рандомизированное двойное слепое контролируемое исследование. J Аффективное расстройство. 2014; 162:43–9.
С., Де Рэдт Р., Бенсенор И.М., Лотуфо П.А., Намюр В. и др. Терапия когнитивного контроля и транскраниальная стимуляция постоянным током при депрессии: рандомизированное двойное слепое контролируемое исследование. J Аффективное расстройство. 2014; 162:43–9.
Артикул КАС пабмед Google ученый
Сегрейв Р.А., Арнольд С., Хой К., Фицджеральд П.Б. Параллельное обучение когнитивному контролю усиливает антидепрессивную эффективность tDCS: пилотное исследование. Мозговой стимул. 2014;7:325–31.
Артикул КАС пабмед Google ученый
Oliveira JF, Zanao TA, Valiengo L, Lotufo PA, Bensenor IM, Fregni F, et al. Резкое улучшение рабочей памяти после tDCS у пациентов без антидепрессантов с большим депрессивным расстройством. Нейроски Летт. 2013; 537: 60–4.
Артикул КАС пабмед Google ученый
 «>
«>Пауэлл Т.Ю., Бунстра Т.В., Мартин Д.М., Лу К.К., Брейкспир М. Модуляция активности коры путем транскраниальной стимуляции постоянным током у пациентов с аффективным расстройством. ПЛОС ОДИН. 2014;9:e98503.
Артикул КАС пабмед ПабМед Центральный Google ученый
Мартин Д.М., Моффа А., Николин С., Беннаби Д., Брунони А.Р., Фланнери В. и др. Когнитивные эффекты лечения транскраниальной стимуляцией постоянным током у пациентов с большим депрессивным расстройством: метаанализ данных отдельных пациентов в рандомизированных плацебо-контролируемых исследованиях. Neurosci Biobehav Rev. 2018; 90:137–45.
Артикул пабмед Google ученый
Вандерхасселт М.А., Брунони А.Р., Лоис Т., Боджио П.С., Де Рэдт Р. Nosce te ipsum-Сократ снова? Контроль мгновенных руминативных самореферентных мыслей путем нейромодуляции эмоциональной рабочей памяти.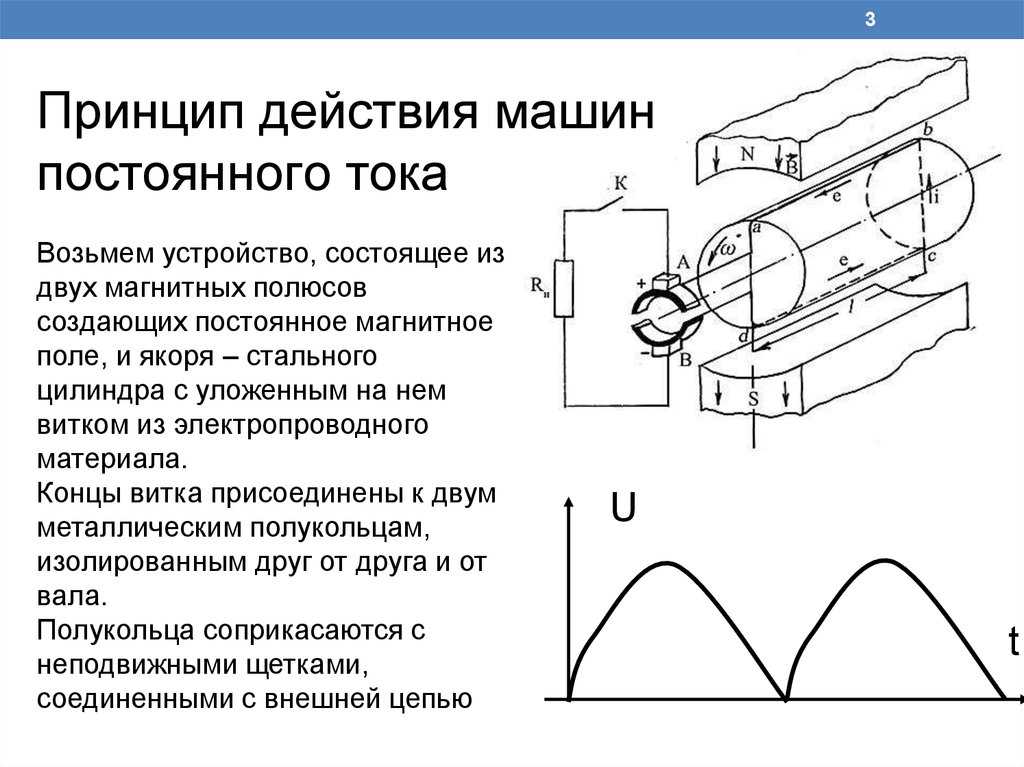 Нейропсихология. 2013;51:2581–9.
Нейропсихология. 2013;51:2581–9.
Артикул пабмед Google ученый
Леш Т.А., Ниендам Т.А., Минценберг М.Дж., Картер К.С. Дефицит когнитивного контроля при шизофрении: механизмы и значение. Нейропсихофармакология. 2011;36:316–38.
Артикул пабмед Google ученый
Mondino M, Jardri R, Suaud-Chagny MF, Saoud M, Poulet E, Brunelin J. Влияние лобно-височной транскраниальной стимуляции постоянным током на слуховые вербальные галлюцинации и функциональную связность левого височно-теменного соединения в состоянии покоя у больных шизофренией. Шизофр Булл. 2016;42:318–26.
Артикул пабмед Google ученый
Brunelin J, Mondino M, Gassab L, Haesebaert F, Gaha L, Suaud-Chagny MF, et al. Изучение транскраниальной стимуляции постоянным током (tDCS) в качестве лечения галлюцинаций при шизофрении. Am J Психиатрия. 2012; 169:719–24.
Am J Психиатрия. 2012; 169:719–24.
Артикул пабмед Google ученый
Фитцджеральд П.Б., Маккуин С., Даскалакис З.Дж., Хой К.Е. Отрицательное пилотное исследование ежедневной бимодальной транскраниальной стимуляции постоянным током при шизофрении. Мозговой стимул. 2014;7:813–6.
Артикул пабмед Google ученый
Кекич М., Бойсен Э., Кэмпбелл И.С., Шмидт У. Систематический обзор клинической эффективности транскраниальной стимуляции постоянным током (tDCS) при психических расстройствах. J Psychiatr Res. 2016;74:70–86.
Артикул пабмед Google ученый
Моффа А.Х., Брунони А.Р., Николин С., Лоо К.К. Транскраниальная стимуляция постоянным током при психических расстройствах: всесторонний обзор. Психиатр Clin North Am. 2018;41:447–63.
Артикул пабмед Google ученый
Френьи Ф., Лигуори П., Фекто С., Ницше М.А., Паскуаль-Леоне А., Боджио П.С. Корковая стимуляция префронтальной коры с помощью транскраниальной стимуляции постоянным током снижает тягу к курению, вызванную сигналом: рандомизированное, ложно-контролируемое исследование. Дж. Клин Психиатрия. 2008; 69: 32–40.
Артикул пабмед Google ученый
Боджио П.С., Султани Н., Фекто С., Мерабет Л., Мекка Т., Паскуаль-Леоне А. и др. Модуляция префронтальной коры с использованием транскраниальной стимуляции постоянного тока снижает тягу к алкоголю: двойное слепое исследование с ложным контролем. Наркотики Алкогольная зависимость. 2008;92:55–60.
Артикул пабмед Google ученый
Батиста Э.К., Клаусс Дж., Фрегни Ф. , Ницше М.А., Накамура-Паласиос Э.М. Рандомизированное плацебо-контролируемое исследование целенаправленной модуляции префронтальной коры с помощью двусторонней tDCS у пациентов с зависимостью от кокаина. Int J Neuropsychopharmacol. 2015;18:pyv066.
, Ницше М.А., Накамура-Паласиос Э.М. Рандомизированное плацебо-контролируемое исследование целенаправленной модуляции префронтальной коры с помощью двусторонней tDCS у пациентов с зависимостью от кокаина. Int J Neuropsychopharmacol. 2015;18:pyv066.
Boggio PS, Zaghi S, Villani AB, Fecteau S, Pascual-Leone A, Fregni F. Модуляция риска у потребителей марихуаны с помощью транскраниальной стимуляции постоянным током (tDCS) дорсолатеральной префронтальной коры (DLPFC). Наркотики Алкогольная зависимость. 2010;112:220–5.
Артикул пабмед Google ученый
Шахбабая А., Голесорхи М., Заманян Б., Эбрахимпур М., Кешвари Ф., Неджати В. и др. Зависящий от состояния эффект транскраниальной стимуляции постоянным током (tDCS) на тягу к метамфетамину. Int J Neuropsychopharmacol. 2014;17:1591–8.
Артикул КАС пабмед Google ученый
 «>
«>da Silva MC, Conti CL, Klauss J, Alves LG, do Nascimento Cavalcante HM, Fregni F, et al. Поведенческие эффекты транскраниальной стимуляции постоянным током (tDCS) индуцировали пластичность дорсолатеральной префронтальной коры при алкогольной зависимости. J Физиол Париж. 2013; 107: 493–502.
Артикул пабмед Google ученый
Берлоу Ю.А., Зандвакили А., Карпентер Л.Л., Филип Н.С. Транскраниальная стимуляция постоянным током при униполярной депрессии и риске лечения эмерджентной мании: обновленный метаанализ. Мозговой стимул. 2019;12:1066–8.
Артикул пабмед Google ученый
Хорват Дж. К., Картер О., Форте Дж. Д. Транскраниальная стимуляция постоянным током: пять важных вопросов, которые мы не обсуждаем (но, вероятно, должны обсуждать). Фронт Сист Нейроци. 2014;8:2.
Артикул пабмед ПабМед Центральный Google ученый
 «>
«>McIntosh RD. Исследовательские отчеты: новый тип статей для коры. кора. 2017; 96:А1–А4.
Артикул Google ученый
Civile C, McLaren R, McLaren IPL. Как мы можем изменить ваше мнение: анодная tDCS в Fp3 изменяет представление человеческого стимула и обучение. Нейропсихология. 2018;119:241–6.
Артикул пабмед Google ученый
Minarik T, Berger B, Althaus L, Bader V, Biebl B, Brotzeller F, et al. Важность размера выборки для воспроизводимости эффектов tDCS. Передний шум нейронов. 2016;10:453.
Артикул пабмед ПабМед Центральный Google ученый
Джалали Р., Миалл Р.С., Галеа Дж.М. Отсутствует стойкий эффект транскраниальной стимуляции мозжечка постоянным током на зрительно-моторную адаптацию. J Нейрофизиол. 2017; 118: 655–65.
Артикул пабмед ПабМед Центральный Google ученый
Вудс А.Дж., Антал А., Биксон М., Боггио П.С., Брунони А.Р., Сельник П. и др. Техническое руководство по tDCS и связанным с ними инструментам неинвазивной стимуляции мозга. Клин Нейрофизиол. 2016; 127:1031–48.
Артикул КАС пабмед Google ученый
Фрид Э.И. 52 симптома большой депрессии: отсутствие содержания перекрываются между семью распространенными шкалами депрессии. J Аффективное расстройство. 2017;208:191–7.
Артикул пабмед Google ученый
Phillips ML, Chase HW, Sheline YI, Etkin A, Almeida JR, Deckersbach T, et al. Выявление предикторов, модераторов и медиаторов реакции на антидепрессанты при большом депрессивном расстройстве: подходы нейровизуализации. Am J Психиатрия. 2015; 172:124–38.
Am J Психиатрия. 2015; 172:124–38.
Артикул пабмед ПабМед Центральный Google ученый
Дункан Дж. Модель адаптивного кодирования нейронной функции в префронтальной коре. Нат Рев Нейроски. 2001; 2: 820–9.
Артикул КАС пабмед Google ученый
Тонони Г., Спорнс О., Эдельман Г.М. Меры вырожденности и избыточности в биологических сетях. Proc Natl Acad Sci USA. 1999;96:3257–62.
Артикул КАС пабмед Google ученый
Харти С., Селла Ф., Коэн Кадош Р. Разум мозга: посредническая и сдерживающая роль нейрофизиологии. Тенденции Cogn Sci. 2017;21:2–5.
Артикул пабмед Google ученый
Радман Т., Рамос Р.Л., Брумберг Дж.С., Биксон М. Роль типа и морфологии клеток коры в подпороговой и надпороговой стимуляции однородным электрическим полем in vitro. Мозговой стимул. 2009;2:215–28. 228 е211-213.
Роль типа и морфологии клеток коры в подпороговой и надпороговой стимуляции однородным электрическим полем in vitro. Мозговой стимул. 2009;2:215–28. 228 е211-213.
Артикул пабмед ПабМед Центральный Google ученый
Карп Дж. О множественности (методологических) миров: оценка аналитической гибкости экспериментов FMRI. Фронтальные нейроски. 2012;6:149.
Артикул пабмед ПабМед Центральный Google ученый
Аль-Кайси А.М., Аль-Ани А., Лу К.К., Пауэлл Т.И., Мартин Д.М., Брейкспир М. и др. Прогнозирование результатов лечения tDCS у пациентов с большим депрессивным расстройством с использованием автоматизированной классификации ЭЭГ. J Аффективное расстройство. 2017; 208: 597–603.
Артикул пабмед Google ученый
Д’Урсо Г.